

La alimentación porcina parece ser una de las estrategias más eficaces para paliar el problema detectado en algunas zonas de España por la elevada cantidad de purines generada. El objetivo de este ensayo fue evaluar el impacto de la reducción del nivel de proteína bruta (PB) de 14,8 a 12,6% en la dieta de finalización de cerdos, mediante la suplementación con aminoácidos esenciales, sobre la productividad, el balance de N, algunos metabolitos séricos, la digestibilidad de nutrientes y la concentración de compuestos olorosos fecales. Además, se realizó una evaluación económica y otra ambiental. Los resultados mostraron que la reducción del contenido de PB en la dieta condujo a una reducción del N excretado, sin penalizar los resultados productivos ni la calidad de la canal pero sí reduciendo el contenido proteico de la carne. Además, esta dieta tuvo menor impacto sobre el medio ambiente que la dieta control.
Introducción, hipótesis y objetivos
España es uno de los líderes mundiales en producción porcina. En 2021, el censo porcino alcanzó 32,7 millones de cabezas y se sacrificaron un total de 56,4 millones de cerdos produciendo alrededor de 5 millones de toneladas de carne de cerdo (FAOSTAT, 2022). Más del 40% de la cría de cerdos se localiza en el norte de nuestro país concentrándose en Aragón y Cataluña (MAPA, 2022). Esa densidad porcina ha generado problemas ambientales y sociales relacionados con la producción de purines. Algunos periódicos afirman que en Cataluña, el 43% de las principales fuentes de agua y los ríos están contaminados por nitratos (NO3) con concentraciones que superan los límites de potabilidad (50mg/L), eutrofización y pérdidas de biodiversidad (IPCC, 2006). Esta contaminación está provocada por un exceso de nitrógeno (N) en los purines que, una vez distribuidos en el campo, contaminan las aguas superficiales y profundas provocando la contaminación por NO3-. Los principales compuestos de N en los purines porcinos son: amoníaco (NH3), óxido nitroso (N2O) y óxido de N (NO3-).
Se han generado varias herramientas de cálculo del contenido de N de las deyecciones en algunas CC.AA., es el caso de la publicada por el Gobierno de Aragón (BOA, 2021). Esta herramienta también regula los criterios y procedimientos para el reconocimiento de la reducción en la excreción de N respecto a los valores estándar mediante la mejora en la alimentación. Los datos obtenidos del balance de N son especialmente útiles en zonas con alto censo, ya que permite una mejor estimación de la composición de los purines y un mejor ajuste de las dosis a aplicar a los cultivos.
Para evitar el exceso de N excretado, se están evaluando varias estrategias. Una de las más efectivas es optimizar el uso de las proteínas en la dieta por parte de los cerdos. La eficiencia de utilización de la proteína depende de la digestibilidad de la fuente de proteína bruta (PB), del contenido de aminoácidos (AA) de la proteína alimentaria y de los requerimientos de AA del cerdo (Van Milgen y Dourmad, 2015). Si la cantidad de AA está en exceso en la dieta, los cerdos la catabolizan produciendo más urea en la orina. Es fundamental que la dieta del cerdo contenga todos los AA esenciales, que son: Lisina (Lys), Metionina (Met), Treonina (Thr), Triptófano (Trp), Fenilalanina (Phe), Histidina (His), Valina (Val), Isoleucina (Ile) y Leucina (Leu). La falta de solo uno de estos AA inhibe la síntesis de proteínas porque los cerdos no tienen la capacidad metabólica para sintetizarlos. Además, la composición de AA varía entre los diferentes componentes del cuerpo. Por ejemplo, la producción de 1kg de tejido magro en un cerdo en crecimiento requiere menos AA que la producción de 1L de leche en una cerda lactante. En teoría, para optimizar la utilización de AA por parte de los cerdos, la cantidad suplementada en la dieta debe ser la misma cantidad de AA necesaria para la síntesis de proteínas.
Así es como surge el concepto de proteína ideal, con un objetivo importante; se refiere a la situación en la que todos los AA esenciales están co-limitando de cara al rendimiento productivo de los animales, por lo que el aporte de AA coincide exactamente con las necesidades de AA. Los requerimientos de AA en la proteína ideal generalmente se expresan en relación con los requerimientos de Lys (Lys=100%). La expresión relativa a la Lys es muy útil desde un punto de vista práctico. La Lys es típicamente el primer AA limitante en las dietas para cerdos. Si los requerimientos para los otros AA se conducen principalmente a las necesidades para la síntesis de proteínas, los requerimientos de estos AA deberían ser relativamente constantes (en relación con Lys). Esto simplifica enormemente la nutrición práctica porcina porque un nutricionista solo tiene que conocer el cambio en el requerimiento de Lys a lo largo del tiempo y combinarlo con un perfil proteico ideal constante (Van Milgen y Dourmad, 2015). La integración óptima se produce cuando los AA esenciales en la dieta alcanzan una cantidad en la que la implementación adicional no tiene un efecto relevante en el criterio de respuesta (por ejemplo, en el crecimiento diario). En esta situación, los cerdos pueden expresar su máximo potencial de crecimiento porque el suministro de nutrientes no limita los rendimientos productivos. Además, la cantidad de proteína depositada y la cantidad de alimento ingerido modifican las necesidades de AA del cerdo durante el crecimiento. La cantidad de proteína depositada podría calcularse en base a: la masa proteica inicial (correlacionada con el peso vivo -PV- inicial), la cantidad de tejido muscular en la ganancia diaria y la precocidad, que indica si los cerdos son de maduración temprana o tardía. La cantidad de tejido magro depositado y su composición en AA establecen las necesidades esenciales de AA de los cerdos en cada fase de crecimiento.
Es bien sabido que, en la fase de finalización, los cerdos depositan menos tejido magro que en otras fases y varios estudios han demostrado que, en esta fase, los cerdos tienen menos requerimientos de PB que en la fase de crecimiento (Prandini et al., 2013). Sin embargo, para garantizar buenos rendimientos y una óptima deposición de tejido magro en el periodo de crecimiento-finalización, se suelen proporcionar dietas con altos niveles de PB y esto implica también mayores excreciones de N (Le Bellego et al., 2001). Algunos autores han demostrado que, si se cubren los requerimientos esenciales de AA y N total, es posible disminuir la concentración de PB al disminuir la inclusión de harina de soja (Wang et al., 2018). Si la proteína de la dieta está restringida en un 2-4%, es fundamental complementar con Lys, Thr, Met y Trp para evitar el empeoramiento del rendimiento productivo de los animales (Kerr et al., 2003).
Millet et al. (2017) mostraron que son necesarios 6,3kg de N para el crecimiento de un lechón de 8 a 110kg PV. El 54% de este N lo pierden los cerdos a través de heces y orina. Para reducir esta excreción, la maximización de la eficiencia de utilización del N es clave, aumentando la cantidad de N retenido por los cerdos para producir carne magra sin aumentar la cantidad de N ingerido a través del alimento. Existen diferentes estrategias para mejorar la utilización de N por parte de los cerdos. Por ejemplo, Cruzen et al. (2013) concluyeron que los cerdos con alta eficiencia alimenticia se caracterizan por una mejor utilización del N debido a una menor regeneración de proteína muscular y, por supuesto, un menor consumo de alimento por kg de tejido magro producido. El ajuste del contenido de AA en la dieta a las necesidades de cada fase de crecimiento de los cerdos es una estrategia exitosa para reducir la excreción de N. De hecho, los problemas de la alimentación convencional en 3 fases para cerdos en crecimiento-cebo implica una disponibilidad limitada de nutrientes en la 1ª fase y un exceso en la última fase (Millet et al., 2017). Cuando aumenta el número de esas fases, el sistema de alimentación cambia a una alimentación multifase, por lo que la cantidad de proteína ingerida se adapta mejor a las necesidades de los cerdos. Pomar et al. (2014) observaron una reducción del 12% en el N excretado y una mejora en la eficiencia de utilización del N mediante el uso de un programa de alimentación de varias fases. Con la mayor disponibilidad de AA cristalinos se ha hecho posible aplicar una gran reducción en el nivel de PB en la dieta. Para lograr una reducción considerable de la PB, sin causar una deficiencia de AA no esenciales, es importante que la relación Lys:proteína digestible ileal estandarizada se mantenga entre 0,065 y 0,068 (Henry y Dourmad, 1993).
Por otro lado, el sistema intensivo de producción porcina tiene un impacto negativo en el cambio climático, no solo por la producción de metano, NH3, N2O y otros gases que afectan al clima, sino también por el gran consumo de alimentos, energía y agua. La fuente más importante de preocupaciones ambientales es la producción de piensos. Debido al origen global de los cultivos utilizados para producir piensos compuestos, este proceso tiene un gran impacto en el medio ambiente. En el caso de Europa, los cereales energéticos (cebada y trigo) provienen, en parte, del norte de Europa, la harina de soja de Brasil, los AA sintéticos de China y Corea y las fuentes minerales de Reino Unido (McAuliffe et al., 2016). El mayor impacto de los piensos se debe al transporte y procesamiento de productos de origen vegetal. Por ejemplo, la implementación de la harina de soja en las dietas de finalización de los cerdos tiene un impacto importante debido a que se procesa y transporta desde Brasil, pero la harina de soja se puede reemplazar en estas dietas por una correcta suplementación de AA sintéticos. Ogino et al. (2013) demostraron que la alimentación con una dieta baja en PB complementada con AA esenciales produjo un 20% menos de gases de efecto invernadero (GEI) y redujo los impactos de acidificación y eutrofización. Además, la gestión del estiércol parecía tener un impacto menor y las emisiones de amoníaco se redujeron probablemente debido al menor pH de los purines. Sin embargo, la dieta baja en PB es más cara desde el punto de vista energético debido al proceso industrial para crear AA cristalinos. Otra forma de reducir el impacto ambiental de la producción porcina es mejorar la eficiencia alimenticia para lograr una mayor producción de carne con la misma cantidad de alimento. Esto podría ser interesante en el caso de una dieta baja en PB para cerdos de finalización. Como se ha mencionado anteriormente, varios autores han demostrado que cerdos alimentados con dietas bajas en PB complementadas con AA esenciales mantienen los mismos rendimientos productivos que cerdos alimentados con dietas tradicionales. Sin embargo, debido a la reducción de la inclusión de harina de soja, la carne de cerdos alimentados con dietas bajas en PB tiene menos efectos negativos sobre el medio ambiente.
La producción intensiva de cerdos se ve afectada por muchos costes que involucran, entre otros, alimentos, mano de obra, capital e instalaciones. El beneficio de una granja porcina es el balance económico entre los costes de producir cerdos y los ingresos por venderlos. Estos costes difieren entre los distintos países europeos pero, en todos ellos, la alimentación es el que más influye. Por lo tanto, una posible estrategia para reducir los gastos de alimentación es modificar la composición de la alimentación, reduciendo la inclusión de PB sin penalizar el rendimiento y la calidad de la carne. En China, cada 10g/kg de reducción de PB se produce una reducción del 1,50% del coste de alimentación (Wang et al., 2018). También hay que tener en cuenta que el precio de los piensos se ve afectado por la fluctuación de los precios de los piensos.
En este contexto, la hipótesis es que, en la fase de finalización del engorde de los cerdos, es posible una reducción de los niveles nutricionales, manteniendo constantes los AA digestibles en relación con el suministro de PB, sin perjudicar la productividad ni la calidad y obteniendo ventajas ambientales.
El objetivo de este ensayo fue estudiar el impacto de la reducción del nivel de PB en la dieta de finalización de cerdos, mediante la suplementación con AA esenciales, sobre los rendimientos productivos, el balance de N, los niveles de metabolitos séricos, la digestibilidad de los nutrientes y la concentración de compuestos olorosos fecales. Además, se realizó un análisis de ciclo de vida (ACV) para evaluar el impacto ambiental de esta estrategia alimentaria.
Material y Métodos
Todos los procedimientos experimentales utilizados en este estudio cumplieron con las directrices españolas para el cuidado y uso de animales en investigación (BOE, 2013), en lo que respecta a la protección de animales utilizados para experimentación y otros fines científicos.
- Animales y dietas experimentales
Se utilizaron un total de 80 cerdos, 50% machos enteros y 50% hembras, de cruce (Landrace x Large White) x Pietrain, en un experimento que duró el período de finalización (35 días). El peso y la edad iniciales fueron 92,7 ± 3,61kg PV y 110 ± 3 días. Al inicio de la prueba, los cerdos fueron pesados y alojados en una nave de ambiente natural, con corrales de 7,5m2 con suelo emparrillado que incluían un bebedero de cazoleta (2L/min de caudal de agua) y un comedero tipo tolva holandesa con alimentación seca. Los animales se distribuyeron en 8 corrales (10 cerdos/corral) por sexo y según 2 dietas en función del nivel proteico de la dieta: control (15% PB) y bajo en PB (13%). Estas dietas fueron formuladas para cumplir con el concepto de proteína ideal. En la dieta baja en PB, la reducción de harina de soja se sustituyó principalmente por cereales (maíz, trigo y cebada) y hubo que añadir AA sintéticos cristalinos para evitar deficiencias de AA (tabla 1).
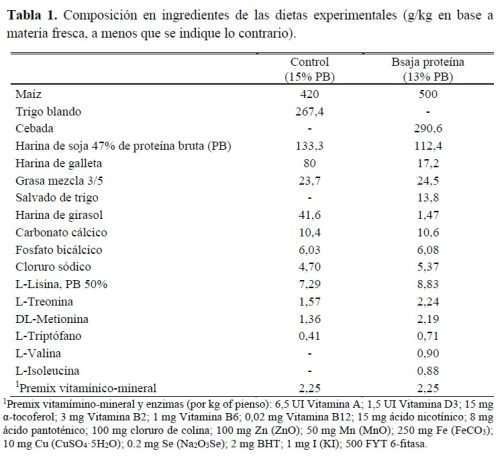
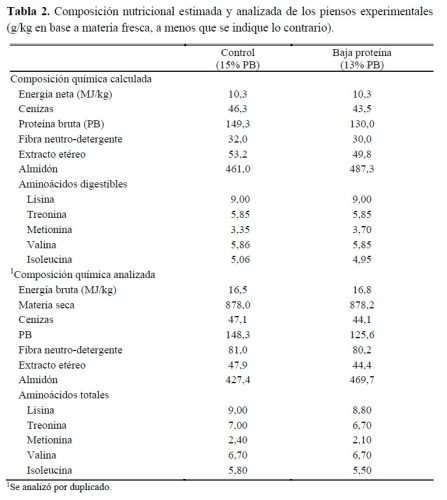
Ambas dietas se formularon para ser isoenergéticas (10,3MJ de energía neta/kg) e isoaminoácidas pero con un contenido de PB más bajo (150 y 130g de PB/kg). Por lo tanto, cada dieta experimental se replicó 4 veces siendo la réplica el corral (2 corrales de machos y 2 corrales de hembras por dieta). Los piensos se proporcionaron ad libitum y en forma granulada durante todo el ensayo. La tabla 2 muestra la composición química calculada y analizada de las dietas evaluadas en el presente experimento.
2. Rendimientos productivos, balance de N balance y muestreos en la granja
Se registró el PV individual de los cerdos los días 0, 15 y 35 del ensayo. Asimismo, se evaluó el consumo de alimento por corral entre esas fechas considerando la oferta de alimento y los rechazos. Estos datos se utilizaron para calcular la ganancia media diaria (GMD), el consumo medio diario de alimento (CMD) y el índice de conversión (IC) por corral para todo el período de finalización.
Con los resultados de GMD, CMD, porcentaje de PB real de las dietas y PV inicial y final de los cerdos se estimó el balance de N en los animales según el método de cálculo de la normativa Regional (ORDEN ARP/225/2019). En este sentido, el N ingerido se calculó a partir del consumo de alimento y su contenido en N; el N retenido con la ganancia de PV; el N excretado por diferencia entre el N ingerido y el N retenido; y finalmente, la eficiencia de N como la relación entre el N retenido y el N consumido, siguiendo la metodología de Gallo et al. (2014).
Se tomaron muestras de los piensos los días 0, 15 y 35 del ensayo y se almacenaron a temperatura ambiente hasta su análisis químico. Los mismos días se tomaron muestras fecales de 3 animales por corral, directamente por estimulación rectal. Las heces de cada corral se mezclaron en un pool para obtener muestras homogéneas y se almacenaron en congelación (-20ºC) hasta los análisis de laboratorio de compuestos volátiles y digestibilidad de nutrientes. También se tomaron muestras de sangre de 3 cerdos de cada corral, en los días 5 y 35 del ensayo (siempre de los mismos animales), mediante venopunción yugular en tubos desechables. Inmediatamente, las muestras de sangre se centrifugaron (1200 x g, 10min) y se almacenaron a -20°C hasta el análisis de los metabolitos séricos.
3. Análisis laboratoriales de muestras de piensos, heces y sangre
Las muestras de pienso fueron analizadas por triplicado, siguiendo los procedimientos de la AOAC (2005), para: materia seca (MS) por el método de secado en horno (934.01), cenizas totales en horno mufla (942.05) y PB mediante el método Kjeldhal (976.05) utilizando un Analizador Kjeldahl MT 2300. El extracto etéreo (EE) se determinó usando tecnología Ankom (Ankom Extraction System XTI5), sin hidrólisis previa, y la fibra neutro-detergente (FND) mediante un analizador ANKOM 220, de acuerdo con lo descrito por Mertens (2002), y se expresó como residuos libres de cenizas. El almidón total se analizó enzimáticamente (Comisión de las Comunidades Europeas, 1999) utilizando un kit comercial (Total Starch Assay Kit K-TSTA07/11 Megazyme). La composición en AA se analizó en un laboratorio externo mediante fuorescencia por HPLC (PNT-M-109). La energía bruta se determinó mediante una bomba calorimétrica.
En las muestras de heces se midió el pH antes del secado a 60°C para determinar el contenido de MS como indicador de la consistencia fecal. Después se analizaron los ácidos grasos volátiles (AGV), mediante cromatografía de gases (utilizando 1g de heces frescas) para relacionar cada dieta con la concentración de compuestos olorosos producidos (acético, propiónico, butírico, isobutírico, valérico e isovalérico). Estos análisis se realizaron de acuerdo con una modificación del método de Schauf et al. (2018). Además, las muestras fecales se utilizaron para calcular la digestibilidad de los nutrientes, siendo analizadas por triplicado para: MS, materia orgánica (cenizas totales), PB, EE y FND, utilizando los mismos procedimientos descritos anteriormente para las dietas. Además, se calculó la digestibilidad del fósforo (P) y de los carbohidratos (CH). Así, a partir de piensos y heces, se determinó el P total mediante espectrofotometría de emisión atómica (ICP-OES, HORIBA Jobin Yvon, familia Activa, con AS-500 Autosampler, HORIBA Scientific) y se estimó el total de CH mediante la siguiente expresión: CH=MS–(PB+EE+Cenizas). El método utilizado para estimar la digestibilidad de los nutrientes fue el basado en cenizas insolubles en ácido clorhídrico (AIA). El análisis de AIA (BOE, 1995) permite determinar el contenido de materia mineral insoluble en ácido tanto en piensos como en heces. Para el cálculo del coeficiente de digestibilidad aparente de los nutrientes se utilizó la siguiente ecuación:
Coeficiente de digestibilidad aparente (%) = 100 [100 (Md/Mh) (Zh/Zd)] donde Zh y Zd son las concentraciones de nutrientes (%) en las heces y en la dieta, respectivamente, y Mh y Md son las concentraciones (%) de AIA en las heces y en la dieta, respectivamente.
El suero de las muestras de sangre se analizó en un laboratorio externo para determinar urea, creatinina, proteínas totales, albúmina, fosfatasa alcalina (FA) y gamma-glutamil transferasa (GGT).
4. Medidas tomadas en la canal y en la carne en el matadero
Cuando los cerdos alcanzaron el peso objetivo de sacrificio (115-117kg PV), se trasladaron al matadero. El ayuno duró aproximadamente 15 horas (sin alimento pero con libre acceso al agua). Los cerdos fueron aturdidos por exposición a CO2 (Butina Aps, 86-88% de concentración media de gas en la atmósfera). A continuación fueron desangrados, escaldados, depilados y eviscerados de acuerdo con los procedimientos comerciales estándar y serrados por la mitad. Antes de la refrigeración, se tomó el peso individualmente de las canales calientes y estos datos se utilizaron para calcular el rendimiento de la canal. De cada canal se estimaron diferentes parámetros utilizando un Autofom III; proporción magra de la canal y peso y porcentaje magro de los 4 cortes principales (jamón, paleta, lomo y panceta), así como la calificación de la canal SEUROP (BOE, 2011), que se basa en el porcentaje de carne magra, y el espesor de grasa subcutánea del jamón. Con los datos de los pesos de estas piezas también se calculó su rendimiento en la canal. El precio por kg de canal se ajustó a la clasificación SEUROP correspondiente siguiendo las normas comerciales.
Tras 72h de refrigeración, las canales se despiezaron. Al final de la línea, de la parte caudal de cada media canal izquierda, se cogió una muestra de lomo (aproximadamente 300 g). Inmediatamente después se tomaron las siguientes medidas de todas las muestras de lomo de cerdo; el pH con un pHmetro portátil equipado con una sonda de penetración (Crison 506), el color subjetivo y marmoleo utilizando la escala manual del National Pork Board (NPB, 2011) y el color objetivo con un espectrofotómetro Konica Minolta CM-700d en el espacio CIELab (CIE, 1978), que proporciona valores de L* (luminosidad), a* (tendencia al rojo) y b * (tendencia al amarillo). El equipo proporcionó dos variables de color más; C*ab que se refiere a la saturación de color (índice cromático) y hab que se refiere al tono (Lebret et al., 2015).
5. Análisis laboratoriales de carne
El estudio de la calidad de la carne se realizó con 40 muestras (5 elegidas al azar de cada corral siendo entonces 10 por dieta y sexo) que se conservaron a ‒20˚C hasta su posterior análisis. Cuando llegó el momento, las muestras se descongelaron durante 24 h a 4˚C. Las pérdidas por cocción y la dureza se midieron por el método descrito por Honikel (1998). En 1º lugar se pesaron una por una, se colocaron en bolsas de plástico individuales y se introdujeron en un baño maría a 75˚C hasta alcanzar los 70˚C (Precisterm, J.P.). Durante la cocción, se controló la temperatura interna de las muestras mediante de un termopar tipo T conectado a un registrador de datos (testo 177-T4, Testo GmbH). Después se enfriaron, se secaron y se pesaron nuevamente. Las pérdidas por cocción se calcularon con el peso antes y tras el cocinado y se expresaron en porcentaje. Para medir la dureza, las muestras cocinadas se cortaron en piezas en forma de prisma con una sección transversal de 100mm2 (10×10) con la dirección de las fibras paralelas a la dimensión larga de al menos 30 mm. Se cortaron un total de 8-10 prismas por muestra de forma perpendicular a la orientación de las fibras musculares, con una cizalla Warner-Bratzler conectada a un texturómetro Instron Universal (Modelo 5543) conectado a un ordenador.
La composición química (contenido en humedad, proteína, cenizas y grasa intramuscular -GIM-) de la carne se analizó según los procedimientos del BOE (1979). Para ello, las muestras fueron previamente picadas. La humedad se determinó mediante secado en horno (Memmert UFE500), cenizas en mufla a 550ºC durante 6h, proteína con un analizador 2300 Kjeltec (Foss Tecator) y GIM mediante un Sistema de Extración ANKOMXT15 (ANKOM Techonology) tras ser hidrolizadas las muestras mediante un equipo de hidrólisis ANKOMHCL.
Finalmente, para el genotipado del gen del Halotano, en 1º lugar, las muestras de carne sin humedad se lisaron añadiendo tampón de lisis y proteinasa K durante 6 h a 56°C. Tras la lisis, el ADN se aisló mediante la adición de fenol y cloroformo, dando lugar a 2 fases; una acuosa que contiene los ácidos nucleicos y una orgánica. Posteriormente, el ADN de la fase acuosa se precipitó con un volumen de isopropanol y se resuspendió en tampón TE. La concentración de ADN se evaluó en un espectrofotómetro Nanodrop-100 y la integridad se determinó mediante electroforesis en gel de agarosa. La mutación puntual C1843T en el gen RYR1 (Fuji et al., 1991) se evaluó mediante RFPL-PCR utilizando la restricción de enzima Hhal.
6. Evaluación del impacto ambiental
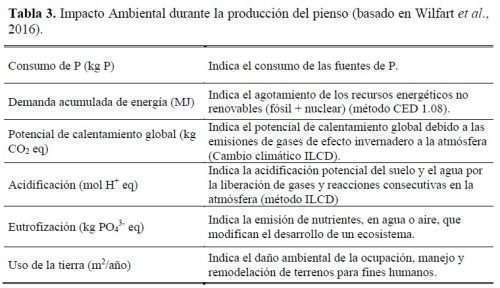
Se realizó una evaluación del impacto ambiental de la producción de los piensos utilizando el método del ACV (Wilfart et al., 2016). Utilizando las tablas INRA ECOALIM, se estimaron los impactos en la producción de cada componente del pienso para varios atributos (tabla 3). La unidad funcional (UF) considerada fue 1kg de pienso.
7. Análisis estadístico
Los datos se analizaron mediante el paquete estadístico JMP Pro14 (SAS Institute Inc. Cary). Los datos de rendimientos productivos, el balance de N, las variables de calidad de la canal y de la carne y los indicadores económicos se analizaron con modelos simples de mínimos cuadrados que incluyeron el tratamiento alimentario (control o bajo en PB), el sexo (machos y hembras) y su interacción como efectos fijos. Las determinaciones fecales (pH, MS, AGV y digestibilidad de nutrientes) se analizaron con el mismo modelo pero considerando también como efectos fijos el día de muestreo (1, 15 o 35) y su interacción con el tratamiento alimentario y el sexo. En estas variables se consideró el corral como unidad experimental (n=4 repeticiones por dieta). Los metabolitos sanguíneos se analizaron con un modelo mixto con medidas repetidas que incluía como efectos fijos el tratamiento alimentario, el sexo y el día de muestreo (5 o 35), así como sus interacciones individuales, y el animal como efecto aleatorio. En este caso, se consideró como unidad experimental al cerdo muestreado (n=12 repeticiones por dieta). Los resultados se expresan como medias corregidas por mínimos cuadrados y sus errores estándar. La separación de medias se realizó con la prueba de Tukey. El nivel de significancia se fijó en 0,05, pero se comentan las tendencias si el nivel de significación está por debajo de 0,10.
Resultados, Discusión y Conclusiones
1. Rendimientos productivos
Los resultados del impacto del nivel de PB en la dieta en la productividad de los cerdos se muestran en la tabla 4. No se detectaron diferencias en el peso final (peso al sacrificio) debido al tratamiento (P>0,10), pero sí una diferencia significativa ( P<0,05) entre el PV de machos y hembras, cuyo valor inicial y final en machos fue de 95,3 y 121,5kg y en hembras de 89,5 y 114,1kg, respectivamente. Esto es lógico ya que eran animales de la misma edad y, a la misma edad, la literatura coincide en que los machos pesan más que las hembras (Latorre et al., 2003; Van den Broeke et al., 2020). Aunque los machos mostraron mayor GMD y mejor IC que las hembras, las diferencias tampoco fueron significativas (P>0,05).
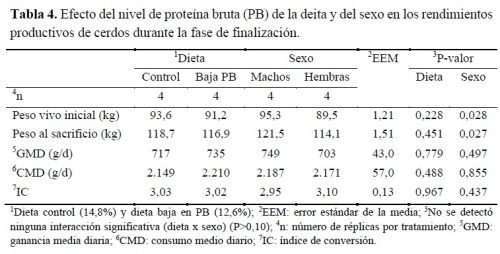
A pesar de las diferencias numéricas observadas en la GMD, el CMD y el IC entre dietas, no fueron significativas en ningún caso (P>0,05), por lo que se puede deducir que esta reducción del contenido de PB en la dieta de cerdos en finalización no supone una penalización de los rendimientos productivos de los animales, lo que confirma los resultados de otros autores (Kerr et al., 2003; Zhang et al., 2016; Wang et al., 2019).
Otros trabajos estudiaron el efecto de la reducción de la PB sin suplementación con AA, obteniendo productividades similares a las obtenidas con dietas estándar; este es el caso del estudio realizado por Tous et al. (2014). Sin embargo, otros (Nyachoti et al., 2003) observaron una disminución significativa en la GMD y en el CMD cuando los cerdos fueron alimentados con dietas bajas en PB, posiblemente debido a una deficiencia en algunos AA como Ile y Val. Cabe señalar que la disminución del contenido de PB en la dieta influye más en la fase de crecimiento que en la fase de finalización de los cerdos. Así lo confirman algunas investigaciones como las de Apple et al. (2017) o Gloaguen et al. (2014). Asimismo, Suárez-Belloch et al. (2016), con una reducción en el contenido de PB en la dieta sin suplementación con AA durante los primeros 45 días del cebo, detectaron inicialmente una reducción en el crecimiento y el consumo de alimento y un empeoramiento en la eficiencia alimentaria; sin embargo, durante el período de finalización, cuando la dieta era común para todos los cerdos, hubo una compensación con mayor tasa de crecimiento y consumo de alimento y con una mejora del IC.
2. Balance de N
La tabla 5 muestra los resultados estimados sobre el balance de N en los cerdos en función del nivel de PB de la dieta y del sexo.
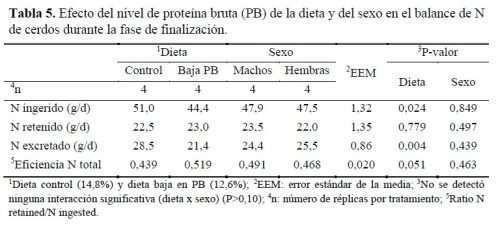
La dieta influyó en el N ingerido (P<0,05), arrojando valores de 51,0 y 44,4g/d para las dietas control y baja en PB, respectivamente. Esta diferencia se atribuye a la composición del pienso, ya que el parámetro se calculó a partir del CM, que fue similar entre dietas, y al contenido de PB de la dieta, que fue notablemente diferente (14,8 vs 12,6%). Por otro lado, el N retenido, calculado a partir de la GMD y el contenido de N muscular, fue similar entre dietas (P>0,05). En cuanto al N excretado, fue significativamente mayor en el caso de los cerdos alimentados con la dieta control que en los cerdos con baja PB (28,5 vs 21,4g/d; P=0,004). Por tanto, los animales alimentados con la dieta baja en PB mostraron una mayor eficiencia en el uso de N (P=0,051), ya que fueron capaces de retener prácticamente el mismo N habiendo ingerido menos.
Finalmente, no se detectaron diferencias entre machos y hembras para ninguna de las variables estudiadas relacionadas con el balance de N (P>0,05), lo que era de esperar ya que tampoco hubo diferencias en los comportamientos productivos. Varios estudios han mostrado resultados similares al presente trabajo. En el de Gallo et al. (2014), no se observó variación en el N retenido cuando se redujo el nivel de PB en la dieta, pero sí se redujo la excreción de N en un 22% con respecto a la dieta control, observándose una tendencia a una mejor eficiencia del N en los animales alimentados con la dieta reducida en PB, independientemente del sexo. Por otro lado, Schiavon et al. (2019) encontraron, en cerdos engordados hasta 145kg PV, que se reducía el contenido de PB y AA manteniendo constantes los contenidos de Lys, Met, Trp y Thr por unidad de PB. Además, Gloaguen et al. (2014) estudiaron el efecto de la reducción de proteína con la suplementación de AA en lechones destetados observando una reducción del N retenido al reducir el contenido de PB de 14,0 a 12,7%. En un estudio realizado por Zhao et al. (2019) se obtuvieron resultados similares; una reducción de la PB del 2 ó 4% y el ajuste con AA esenciales condujo a una reducción de la excreción de N del 16,9 y el 31,9%, respectivamente, en comparación con la dieta rica en PB.
3. Composición de las heces, pH y producción de compuestos olorosos
No se detectó ningún efecto del sexo sobre la MS, el pH ni la concentración de AGV en las heces de los cerdos y, por lo tanto, en la tabla 6 solo se presenta la influencia de la reducción de la PB en la dieta y el día del muestreo. Las heces mostraron una tendencia a estar más secas cuando los cerdos recibieron una dieta baja en PB (P=0,10) y también resultaron más secas los días 15 y 35 del ensayo que el día 0 (P=0,001). Podría tener implicaciones interesantes sobre la cantidad total de estiércol producido por cerdos en periodo de finalización; las heces con alto contenido de MS pierden menos agua y por lo tanto el volumen total de estiércol sería menor. Además, un estiércol más seco podría tener efectos positivos en la separación de las fases del estiércol en las fases de finalización, así como en la reducción de los costes de esparcimiento del estiércol; los resultados de la separación son más efectivos cuando aumenta la MS debido al probable aumento de la materia sólida extraída en comparación con un estiércol más líquido.
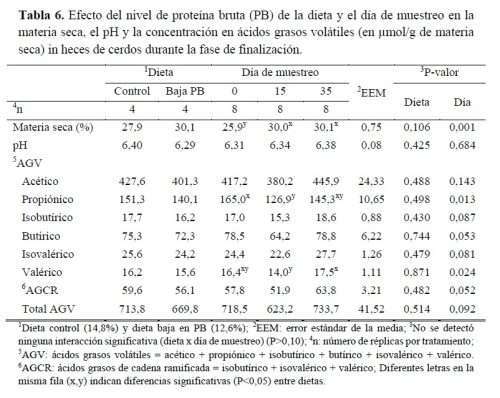
El pH de las heces no se vio afectado por el nivel de PB en la dieta ni por el día de muestreo. Sin embargo, Morazán et al. (2015) registraron un pH más bajo del purín (mezcla de heces y orina) de cerdos alimentados con una dieta menos proteica que los alimentados con dietas más proteicas. El N urinario podría ser el responsable de los diferentes pH de los purines pero no en las heces.
La emisión de olores del engorde de cerdos se debe a la generación de grandes cantidades de orina y heces y a la fermentación anaeróbica de este estiércol durante el almacenamiento. El estiércol contiene residuos de alimentos, nutrientes no digeridos y bacterias muertas del tracto gastrointestinal de los cerdos que son fermentados anaeróbicamente por la población microbiana en el purín y originan cuatro grupos principales de compuestos activos del olor (Le et al., 2007). Varios estudios coinciden en una reducción importante de los compuestos olorosos fecales al disminuir la PB de la dieta. Los carbohidratos se catabolizan a ácidos grasos de cadena corta (AGCC), CO2, H2 y CH4 y las proteínas son los precursores de los AGCC, los ácidos grasos de cadena ramificada (AGCR), los fenoles, los indoles, el azufre, el amoníaco y las aminas (Jensen y Jørgenses, 1994).
En el presente trabajo, la reducción de PB en la dieta no influyó en ningún AGV y el día de muestreo solo afectó a algunos AGV (ácidos propiónico, butírico, isovalérico y valérico), cuya concentración en las heces disminuyó inesperadamente en la mitad del ensayo aumentando el último día (propiónico e isovalérico P<0,05 y butírico y valérico P<0,10). Aunque las diferencias fueron solo numéricas, hay que señalar que la disminución de la suplementación con PB del 14,8 al 12,6 % provocó una ligera reducción lineal en el contenido total de AGV (714 a 670 µmol/g heces de MS, respectivamente; P>0,05). Esto sugiere que el estiércol producido por cerdos alimentados con una dieta baja en PB resultaría menos concentrado en compuestos malolientes (-6,2% total de AGV). Hayes et al. (2004) encontraron que dietas con 16 y 13% de PB produjeron una reducción en la emisión de olores de 31-33% en comparación con una dieta con un 19% de PB. En otro experimento, Cho et al. (2015) informaron que la reducción del nivel de PB del 20 al 15% generó una producción más baja de fenoles, indoles, SCFA y BCFA en un 35-40%. La razón de la falta de efecto en el presente estudio podría ser la siguiente. En ambos estudios, la dieta control incluía un nivel de PB más alto que la dieta control testada por nosotros (20 y 19 vs 14,8%, respectivamente), hubo un mayor desperdicio de AA en los tratamientos con PB alta, siendo la causa de los AGV obtenidos. En nuestro caso, las dietas estaban bien balanceadas en energía y AA totales (al complementar con AA para cumplir con los requisitos totales de AA), se podría producir un menor desperdicio de AA y se esperaría menos fermentación de carbohidratos y AA en el tracto gastrointestinal.
4. Digestibilidad de los nutrientes
No se encontraron interacciones significativas entre tratamiento alimentario, sexo y día de muestreo de heces para ningún coeficiente aparente de digestibilidad de ningún nutriente (P>0,10), por lo que solo se muestran en la tabla 7 los efectos fijos.
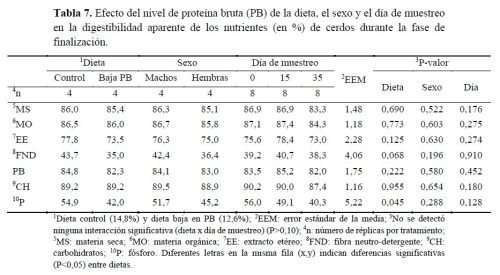
La digestibilidad de los nutrientes no se vio afectada por el tipo de pienso excepto en el caso del P que fue menor (P<0,05) con la dieta baja en PB (54,9 y 42,0% para la dieta control y la baja en PB, respectivamente). Además, se observó una tendencia (P=0,068) a una reducción en la digestibilidad de la FND con la dieta baja en PB (43,7 y 35,0%, respectivamente). El sexo de los animales no influyó en la digestibilidad de ningún nutriente (P>0,05).
Estos resultados concuerdan con el estudio de Wang et al. (2019), donde no se observó efecto en la digestibilidad de la PB ni del EE al reducir la PB en la dieta de cerdos en finalización, aunque los cerdos alimentados con baja PB tendieron a presentar menor digestibilidad de la MS. Sin embargo, algunos autores (Esteves et al., 2021) sí han observado un aumento en la retención de N y en la absorción de P al reducir el contenido de PB en la dieta de 18,2 a 15,2%, manteniendo el suministro de AA esenciales en cerdos de 30 a 50kg PV. En ingredientes de origen vegetal, especialmente en cereales, entre el 60-80% del P total es fítico que está ligado a estructuras complejas como el ácido fítico limitando su disponibilidad en monogástricos (Capelletti et al., 2012). Una posible explicación a los resultados de mayor digestibilidad de P obtenidos por Esteves et al. (2021) podría ser la menor inclusión de harina de soja en estas dietas, cuya concentración de P fítico (0,36%) es mayor en relación a la cantidad presente en el maíz (0,18%), generando una mayor disponibilidad de P. Por otro lado, en el presente estudio se utilizó cebada como fuente proteica y energética en la dieta baja en PB, mientras que en la dieta control se utilizó trigo; en este sentido, Schemmer et al. (2020) indicaron que la digestibilidad del P es mayor cuando se utiliza trigo como cereal de invierno en lugar de cebada (59,4 vs 44,4%), lo que puede deberse al mayor contenido de fibra de la cebada (tanto FND como fibra ácido-detergente), lo que hace que el P se encuentre en el fitato menos accesibles y los CH estructurales se digieren menos.
5. Metabolitos séricos
Los resultados relativos a las concentraciones séricas de urea y creatinina se muestran en la tabla 8. Los niveles de urea se vieron fuertemente afectados por todos los factores estudiados. Se detectaron 2 interacciones significativas; uno entre el nivel de PB de la dieta y el sexo (P<0,01) y otro entre el sexo y el día de muestreo (P<0,001). El 1º implica que, mientras la concentración de urea en machos enteros se redujo drásticamente al alimentarlos con la dieta baja en PB comparado con la dieta control, en las hembras el nivel fue similar, independientemente de la dieta alimentada (Figura 1), y similar al de los machos alimentados con la dieta control. El 2º implica que no hubo diferencia en los niveles de urea entre machos y hembras al comienzo de la prueba pero, al final, las hembras tenían una mayor concentración de urea. Esto puede indicar una capacidad de acumulación de carne magra diferente entre sexos. Abeni et al. (2018) demostraron que los niveles de urea en plasma eran más bajos en las líneas genéticas de cerdos que depositan más tejido magro que aquellas líneas genéticas con más grasa en su ganancia de peso. Esta idea sería válida también para machos y hembras. De hecho, se ha publicado que las canales de hembras son más grasas que las canales de machos enteros, caracterizadas por un alto porcentaje de carne magra (Van de Broeke et al., 2019). La mayor capacidad de deposición de magro de los machos enteros limitaría el desperdicio de AA. Además, hay que señalar que la pubertad es más temprana en las hembras que en los machos (las hembras podrían alcanzarla al principio del ensayo y los machos cerca del final) y es determinante para las necesidades de nutrientes. De hecho, según el NRC (2012), las diferencias de crecimiento entre sexos son más destacadas entre 100 y 135kg PV, cuando sería necesario adaptar la composición de AA de la dieta en machos enteros y nulíparas.
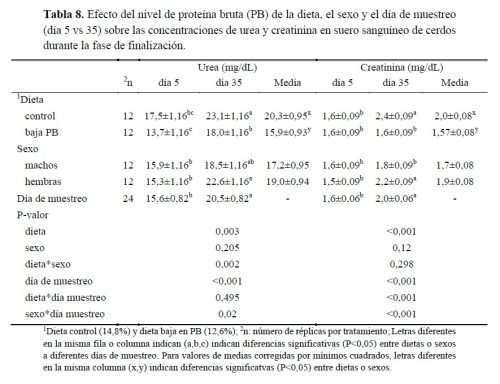
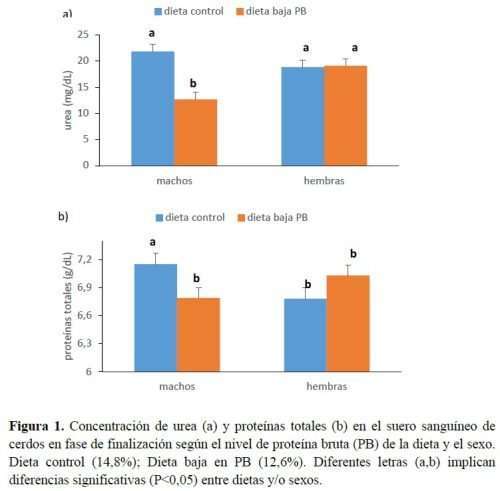
En cerdos alimentados con dietas hiperproteicas, el exceso de PB o AA desequilibrados se transforma en urea en el hígado, y representa un gasto y pérdidas energéticas. Cuando aumenta la concentración de urea en la sangre se excreta más N en la orina y en las heces. La urea se produce por el catabolismo del AA en los cerdos. El N sérico en orina se utiliza como marcador para evaluar la calidad de las proteínas de la dieta y la función renal y hepática. Ma et al. (2019) estudiaron la reducción del contenido de PB en la dieta de cerdos en crecimiento y observaron, como consecuencia, una reducción en el nivel de N en la orina. Este estudio mostró que la dieta baja en PB redujo el nivel de urea sérica, lo que indicaría que una cantidad mínima de AA era catabolizada con esos alimentos y por lo tanto el suministro de PB cumplió con los requerimientos nutricionales de los animales. Esto coincide con estudios anteriores que demuestran que la suplementación con AA esenciales en dietas bajas en PB produce niveles bajos de este metabolito debido a un exceso mínimo de AA para la síntesis de proteínas y una mayor eficiencia del metabolismo de las proteínas (Wang et al., 2019).
Las proteínas séricas totales siguieron una tendencia similar a la de la urea (Figura 1), lo que refleja una menor disponibilidad de la reserva total de AA como sustratos en los machos enteros alimentados con una dieta baja en PB.
La albúmina es una gran fracción de proteína sérica que constituye el 35-50% de la proteína sérica total (Kaneko, 1997). Está implicada en diferentes funciones, como el mantenimiento de la homeostasis, el transporte de sustancias, actúa como eliminadora de radicales libres, regula la presión osmótica del plasma y es la principal fuente de AA que los animales pueden utilizar para satisfacer sus necesidades (Hankins, 2006). Sin embargo, en el estudio actual, la concentración de albúmina no se vio afectada por la dieta o el sexo (P>0,05), que aumentó desde el día 5 al 35 del ensayo (4,3 frente a 4,4 ± 0,05mg/dl, P<0,05).
La creatinina se origina por la transformación de creatina que ocurre principalmente en las células del músculo esquelético; de hecho, el 95-98% de la creatina se almacena dentro del músculo esquelético. La concentración de creatinina sérica está correlacionada con la cantidad de creatina y fosfocreatina en el cuerpo y, en consecuencia, con la cantidad total de masa muscular (Janicki y Buzala, 2013). Se detectaron dos interacciones significativas en la concentración de creatinina; aunque al inicio del estudio el nivel fue similar para todos los animales, los machos mostraron un nivel más bajo al sacrificio que las hembras (P<0,001 para sexo x día de muestreo; tabla 8) y fue menor en la dieta baja en PB que en el control dieta (P<0,001 para dieta x día de muestreo). Esta diferencia entre dietas no concuerda con otros trabajos (Fernández-Fígares et al., 2007; Abeni et al., 2018) en los que la reducción de la PB en la dieta no afectó los niveles de creatinina en cerdos en crecimiento y finalización. En nuestro estudio, la menor concentración de creatinina observada en la dieta baja en PB podría estar relacionada con ligeras diferencias en regeneración muscular, ya que el contenido de proteína de la carne también disminuyó
cuando los cerdos fueron alimentados con la dieta baja en PB (datos que se muestran en la siguiente sección). De hecho, otros autores han demostrado que cerdos en crecimiento alimentados con una dieta baja en proteína tenían un área muscular del lomo reducida (Wang et al., 2018). En base a eso, la cantidad reducida de creatinina detectada en cerdos con dieta baja en PB puede estar relacionada con un contenido de músculo esquelético total potencialmente más bajo. Sin embargo, esta reducción puede ser pequeña ya que no se observaron diferencias en los rendimientos productivos entre las dietas estudiadas. Los resultados sobre las concentraciones de urea, albúmina y creatinina se muestran en la tabla 8.
El análisis de las enzimas fosfatasa alcalina sérica (FAS) y gamma-glutamiltransferasa (GGT) puede ser valioso para evaluar los cambios metabólicos durante el crecimiento de los cerdos o como resultado de un proceso de enfermedad subclínica. Se estudiaron las concentraciones séricas de ambas por su posible correlación con la lesión hepatobiliar y, además, se estudió la FAS para evaluar la actividad osteoblástica. En este estudio, y de acuerdo con Abeni et al. (2018), la disminución del contenido de PB en la dieta no tuvo ningún efecto significativo en ninguno de estos parámetros (datos no mostrados). La actividad enzimática de FAS disminuyó del día 5 al 35 del ensayo (253 frente a 202 ± 13,1 UI/L, P<0,01) mientras que la de GGT no difirió (27,7 frente a 26,0 ± 2,12 UI/L, P>0,05).
6. Calidad de la canal y de la carne
Como se ha mencionado anteriormente, el peso al sacrificio fue mayor en los machos (tabla 9), lo que implicó valores de peso de canal más altos en este sexo (P=0,003). Sin embargo, el rendimiento de la canal fue mayor en las hembras (79,1 vs 77,8%; P=0,002), posiblemente como resultado de un mayor peso visceral y testicular en los machos. Aunque los machos presentaron mayor porcentaje de magro en la canal (P<0,0001), las hembras tuvieron mayor proporción de jamón (P=0,039), lomo (P=0,001), paleta (en este caso fue una tendencia, P=0,091) y panceta (P<0,0001) en la canal.

No se observaron diferencias atribuibles al efecto de la dieta en cuanto al porcentaje de magro, al igual que en el estudio de Salazar-Villanea et al. (2020) en cerdos de finalización al reducir el contenido de proteína en la dieta suplementada con Met, Thr y Trp. La reducción de PB en el alimento tampoco acarreó diferencias en el rendimiento de las piezas estudiadas (P>0,05). Wang et al. (2019) probaron el efecto de la reducción proteica y la suplementación con AA en cerdos durante diferentes fases del crecimiento y no detectaron influencia en los parámetros de calidad de la canal (peso al sacrificio, peso de la canal, rendimiento del lomo) o variables de calidad de la carne (color, pH, pérdidas de agua o composición química). Sin embargo, Tejeda et al. (2020) detectaron en el engorde de cerdos ibéricos algunas similitudes con el presente trabajo, al observar rendimientos similares de paleta y jamón con la dieta limitada en PB, también en concordancia con el trabajo de Tous et al. (2014), pero menores rendimientos en canal y lomo, aunque hay que señalar que en dicho estudio no se realizó suplementación con AA en la dieta poco proteica.
La medición del grosor de la grasa dorsal se acepta como una medida importante de la calidad de la canal, que está relacionada con el rendimiento magro; esta es la razón por la que se incluye en todos los esquemas de calificación. Una posible consecuencia al reducir la PB de la dieta porcina es el aumento de la profundidad del tocino dorsal de la canal, siendo deseable un valor alto en el caso de los animales destinadas a la elaboración de embutidos y productos curados (>115kg PV) pero un valor bajo en animales destinados a la venta de carne fresca (<115kg PV), que sería el caso en este ensayo. Varios estudios recogen que, al reducir la PB de la dieta en cerdos de finalización sin suplementación con AA, aumentó el espesor de la grasa subcutánea (Wood et al. 2013; Morazán et al. 2015; Suárez-Belloch et al., 2016), sin embargo, en el presente estudio, y de acuerdo con los realizados por Schiavon et al. (2019) y Wang et al. (2019), donde se mantuvo el aporte de AA esenciales reduciendo el contenido de PB, no se observó efecto de la dieta en este parámetro. Por otro lado, se observaron diferencias en dicha variable debido al sexo, mostrando los machos valores inferiores a las hembras (10,1 vs 11,1 mm, P<0,05), lo que confirma los resultados encontrados por Gispert et al. (2010) debido a que los machos tienen un mayor potencial de retención proteica que las hembras.
El pH también es un atributo determinante de la calidad de la carne, ya que refleja la formación de ácido láctico al reducirse el glucógeno muscular, e influye directamente en las características fisicoquímicas. En el presente trabajo no se observaron diferencias en el pH final debidas a la dieta o al sexo, ni tampoco en el color subjetivo o el veteado. Un aspecto genético a tener en cuenta en relación con el pH y el color de la carne es el gen del halotano o gen RYR1 (receptor de rianodina que regula la liberación de calcio en el músculo esquelético). La proporción de animales portadores del gen del halotano en el grupo alimentado con la dieta control fue del 80% y en el que recibió la dieta baja en PB fue del 70%, pero esta diferencia no fue significativa (P>0,05). Asimismo, por sexos, el porcentaje de animales portadores del gen halotano fue del 80 y 70% para machos y hembras, respectivamente (P>0,05). Esto garantiza que los resultados obtenidos de la calidad de la carne no se ven influenciados por diferencias en las proporciones del gen halotano.
A pesar de la falta de efecto encontrado en el color subjetivo, algunos de los parámetros indicativos del color medido objetivamente sí mostraron influencia del sexo; se observó un mayor enrojecimiento (a*) en la carne de los machos (P<0,05), así como una mayor tendencia al amarillo (b*) en el mismo sexo (P=0,054), de acuerdo con Gispert et al. (2010). El índice de saturación (C*ab) también fue diferente entre sexos, siendo mayor en los machos (P<0,05), pero el ángulo de tonalidad no fue diferente entre los sexos estudiados.
Aunque en el presente estudio no se podrían atribuir diferencias de color a la dieta, algunos autores han encontrado un aumento del tono amarillo al reducir la PB del 17 al 15% (manteniendo 8,5g Lys/kg de pienso) (Alonso et al., 2010). Además, Gallo et al. (2016), con jamones curados y aplicando una reducción progresiva de los contenidos de PB y Lys, también detectaron piezas más pálidas en las hembras, así como menos veteado, en comparación con los machos.
7. Composición química de la carne de cerdo
En el presente estudio se observó una tendencia a mayores pérdidas por cocinado en los animales alimentados con la dieta baja en PB (17,9 vs 15,8%, P=0.054) (tabla 10), aspecto que podría estar relacionado con los valores finales de pH, ya que a medida que se se produce un alejamiento del punto isoeléctrico de las proteínas (5–5,5) hay más cargas positivas y negativas que no permiten la retención de agua, provocando una disminución de la capacidad de retención y, por tanto, mayores pérdidas por cocción. Puede estar relacionado con los resultados observados en el pH final entre dietas, aunque las diferencias no fueron significativas sí fueron numéricamente mayores en la dieta baja en PB (6,02 vs 5,72). Suárez-Belloch et al. (2016) encontraron que las pérdidas por cocinado aumentaron linealmente a medida que la dieta reducía la PB de 21,6 a 13,5%. En ese estudio también observaron valores de dureza más bajos con el contenido proteico reducido, observándose un valor de dureza más bajo con la dieta menos proteica tanto en machos como en hembras, a diferencia del presente estudio donde no se encontraron diferencias en dureza entre sexos o dietas, de forma análoga a los resultados obtenidos en el estudio realizado por Apple et al. (2017). Sin embargo, esta tendencia también podría estar relacionada con el mayor contenido de humedad observado en la carne de los animales del tratamiento bajo en PB (72,6 vs 73,4%, para la dieta control y baja en PB, respectivamente; P=0,002), que en a su vez fue compensado por un menor contenido de humedad en la carne de esos cerdos (24,1 vs 23,2%, respectivamente; P<0,0001) a pesar de la menor retención (numérica) de N observada en este grupo.
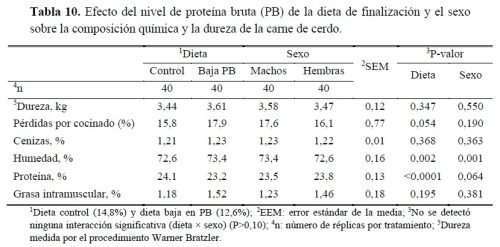
Por otro lado, la carne de los machos mostró mayor porcentaje de humedad (73,4 vs 72,58%, P=0,001) y tendió a presentar mayor contenido en proteína (23,8 vs 23,5%, P=0,064) que la de las hembras. El contenido en GIM o veteado (marmoleado en inglés) es el depósito adiposo que se encuentra asociado a la membrana de los haces musculares (intercelular) o en gotas en las fibras musculares (intracelular). La cantidad y calidad de esta infiltración de grasa son elementos relacionados con el sabor, aroma y terneza de la carne de cerdo. En el presente estudio, el efecto del sexo tampoco afectó el contenido en GIM. Sin embargo, cabe destacar los resultados obtenidos para esta variable al reducir el contenido proteico de la dieta, ya que, a pesar de observarse un contenido graso numéricamente superior en los animales alimentados con la dieta baja en PB (1,52 vs 1,18%), dicha diferencia no fue significativa (P>0,05). Estos resultados difieren con algunos estudios, como el de Tejeda et al. (2020), donde se describe un aumento de la GIM al reducir el contenido proteico de la dieta en cerdos ibéricos, o en el de Wood et al. (2013) quienes obtuvieron mayor contenido en GIM en el lomo. Por otro lado, Alonso et al. (2010) observaron que al reducir la PB del 17 a 15%, manteniendo 8,5g Lys/kg, aumentó la GIM de la carne, sin afectar negativamente las pérdidas por goteo de la misma.
La ausencia de diferencias en el contenido de GIM entre tratamientos alimentarios está en consonancia con los espesores similares de grasa dorsal (grasa subcutánea) detectados en el jamón de cerdos alimentados con ambas dietas. Asimismo, el mayor espesor de grasa en el jamón de las hembras, en comparación con los machos enteros, no se tradujo en una diferencia significativa en el contenido de GIM en la carne.
8. Impacto ambiental
Las 2 dietas diferían en el nivel de inclusión de algunas fuentes de CH y proteínas y se complementaron con diferentes niveles de AA esenciales cristalinos. Los AA cristalinos se sintetizan principalmente en fábricas donde se producen por fermentación bacteriana a partir de azúcar (por ejemplo, jarabe de caña de azúcar) dentro de tanques con condiciones ambientales controladas (pH, temperatura). Posteriormente, los AA se recuperan de las soluciones fermentadas y se cristalizan.
Los efectos de las dietas experimentales sobre el potencial de calentamiento global (PCG) estimado, el uso del suelo y la demanda energética acumulada se muestran en la Figura 2. El mayor impacto se observó en la última variable. Cuando aumentó el contenido de AA en la dieta, aumentó la cantidad de energía consumida por kg de alimento. La energía necesaria para producir 1t de la dieta baja en PB fue un 14% superior a la necesaria para elaborar la dieta control (5.437 MJ/t). Los AA cristalinos cuya fabricación requiere más energía fueron Trp y Val. Alrededor del 22% de la energía total se gastó para producir la suplementación de AA en la dieta de control y esa proporción aumentó al 31% en la dieta baja en PB; en el pienso con PB reducida, un tercio de la energía total consumida para su fabricación puede atribuirse a la elaboración de la suplementación de AA.
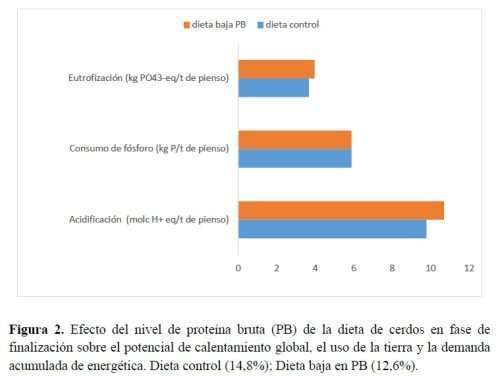
La evaluación del PCG reveló una diferencia mínima entre los dos tratamientos alimentarios. La dieta baja en PB resultó un 5% más impactante que la dieta control, lo que es comprensible dada la enorme huella de carbono que caracteriza la producción industrial de AA (4,9-12,6 t/CO2/t-AA) (Nakamura e Itsubo, 2019). Se podría obtener un efecto positivo en el PCG al reducir el nivel de fuentes proteicas solo si se complementara con una cantidad baja de AA cristalinos. En su estudio, Berton et al. (2018) alimentaron con una dieta baja en proteínas a machos castrados pesados y redujeron en casi un 20% la PB y la Lys digestible en comparación con la dieta convencional (sin equilibrar con AA cristalinos). Estos autores estudiaron cómo las diferentes dietas influyeron en el PCG, la acidificación y la eutrofización mediante el uso de una evaluación de análisis de ciclo de vida. Los resultados evidenciaron que la producción de la dieta baja en PB redujo todos los valores de las categorías de impacto. En nuestro experimento, cuando la suplementación total de AA alcanzó el 1,5% del total de ingredientes en la dieta baja en PB, la emisión de gases del PCG superó los efectos positivos obtenidos por la modulación de la materia prima. Finalmente, la cantidad de suelo consumido/año se elevaba a medida que aumentaba la suplementación con AA pero las diferencias no eran relevantes. Investigaciones recientes han demostrado que el aumento del PCG y los impactos del uso de la tierra durante la fabricación de alimentos con dietas bajas en PB se contrarrestan con menores impactos en la puerta de la granja (por kg de peso corporal del cerdo) (Esteves et al., 2021).
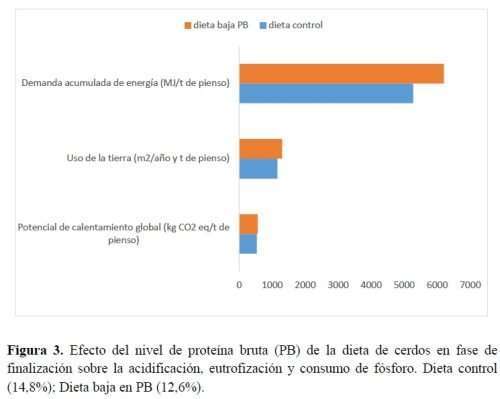
Los efectos de las dietas experimentales sobre la acidificación, la eutrofización y el consumo de P se muestran en la Figura 3. La dieta baja en PB tuvo peor efecto sobre la acidificación y la eutrofización que la dieta control. Estos efectos contaminantes están otra vez relacionados con el aumento de la suplementación con AA que desequilibran los efectos positivos relacionados con la reducción de la inclusión de harina de soja. De hecho, en la dieta baja en PB, debido a la modulación de los ingredientes crudos, H+ y PO43- se redujeron respectivamente en un 7% y un 4% en la dieta baja en PB. Sin embargo, al mismo tiempo, el aumento de la suplementación con AA intensificó los efectos de acidificación y eutrofización en un 50% y un 49% en la dieta baja en PB en comparación con la dieta control. El consumo de P resultó similar entre los dos tratamientos alimentarios y solo fue afectado por la cantidad de fosfato bicálcico agregado a la dieta.
9. Conclusiones
Bajo nuestras condiciones experimentales, podemos concluir que:
- Los rendimientos productivos no se vieron afectados por la reducción de PB de 14,8 a 12,6% en la dieta de finalización de los cerdos y esto además fue independiente del sexo (machos o hembras). Por tanto, la suplementación con aminoácidos sintéticos (Lys, Met, Thr, Met, Trp y Val) cubrió las necesidades nutricionales de los animales.
- La reducción de PB en la dieta provocó una reducción tanto de la ingesta como de la excreción de N en los cerdos, independientemente del sexo, lo que reduciría el impacto de acidificación de la granja.
- La digestibilidad de la mayoría de los nutrientes no se vio afectada por los efectos del sexo o el día de muestreo (edad del animal); sin embargo, la digestibilidad de la fibra neutro-detergente y del fósforo se redujo con la dieta baja en PB, pudiendo comprometer el impacto de eutrofización del medio.
- Los parámetros relacionados con la calidad de la canal y la calidad tecnológica de la carne se vieron influenciados principalmente por el sexo; las hembras presentaron mayor rendimiento de la canal y de las piezas principales (jamón, lomo, paleta y panceta) que los machos, resultando también más grasas.
- La dieta con menor PB afectó a la calidad nutricional de la carne de cerdo disminuyendo el contenido proteico y aumentando el contenido en humedad. El sexo también influyó en estos parámetros, mostrando la carne de las hembras más proteína y menos humedad que la de los machos.
- El nivel de urea fue el metabolito sérico más afectado, siendo fuertemente reducido con una dieta baja en PB al final del experimento. Además, fue menor en los machos que tomaron la dieta baja en PB que en los tomaron la control, pero esta diferencia no se detectó en las hembras. Los niveles de urea en suero también fueron similares entre sexos al comienzo del ensayo, pero más altos en las hembras al final. Esto implica que la dieta control generaría una mayor excreción de N y que sería aconsejable proporcionar una composición de pienso diferente a machos y hembras para reducir la contaminación por N en el purín.
- Las heces estaban más secas con la dieta baja en PB y también a medida que los cerdos aumentaban de peso. La composición del alimento no afectó a ningún ácido graso volátil, aunque se observaron numéricamente menos AGV y AGCM con la dieta con menor PB.
- Según el Análisis de Ciclo de Vida de los piensos, la dieta baja en PB sería la más contaminante, principalmente por el gran impacto de la producción industrial de aminoácidos, que superan el beneficio relacionado con la reducción de fuentes proteicas. Esta diferencia se podría compensar por la menor excreción de nitrógeno de los animales, que permitiría reducir globalmente los atributos de acidificación y necesidad de tierra de la granja.
Autores
- Javier Álvarez Rodríguez. Universidad de Lleida.
- Mª Ángeles Latorre Górriz. Universidad de Zaragoza.
Deja un comentario