


Las características más significativas de la producción de cerdo ibérico (50% Duroc) en categoría de “Cebo de Campo” (BOE, 2014) son las propias de la cría del cerdo tradicional en el suroeste de la península ibérica, en la que se utiliza una raza de cerdo local criado en condiciones de semilibertad en un ciclo largo para producir productos cárnicos de elevada calidad. Estos cerdos se distinguen por su contenido en grasa intramuscular (GIM) y perfil de ácidos graso (AG), principales factores que influyen en la aceptación de un producto cárnico por parte del consumidor (Wood et al., 2008). La raza utilizada procede del cruce al 50% de un macho Duroc y una hembra ibérica. Respecto a otras razas comerciales, destaca por su bajo potencial de crecimiento, elevado perfil adipogénico (come más de lo que necesita, principalmente de la raza ibérica) y potencial contenido en GIM. El sistema de producción utilizado proporciona a los animales suficiente espacio y espacio al aire libre en tierra para caminar y desarrollar un ciclo largo, con una edad y peso vivo (PV) al sacrificio objetivos de 12 meses y 160kg, respectivamente. Este sistema requiere de un programa de alimentación específico que evite el engrasamiento precoz y que tenga en cuenta, por un lado, las necesidades energéticas de mantenimiento extra por las condiciones particulares de alojamiento (ejercicio, ambiente, etc.), y por otro el ajuste de niveles de factores nutricionales y alimenticios que afectan a la composición de AG como son los de energía, proteína, fuentes grasas de los piensos (Essén-Gustavsson et al., 1994; Ninoles et al., 2007) y restricción alimentaria (Daza et al., 2007a). Para ello, se divide en fases a lo largo del crecimiento del animal que combinan piensos de diferente composición y sus diferentes niveles de aporte (racionada o ad libitum), que evitan el engrasamiento precoz, desarrollan un crecimiento compensatorio y dirigen la composición de AG de la grasa de la canal, mediante el control de su síntesis endógena y la acumulación directa (Morel et al., 2006), favoreciendo la deposición de niveles altos de AG omega-3 y oleico. Esta composición grasa de la canal es especialmente sensible a la oxidación y requiere de la incorporación de antioxidantes en el pienso para reducir su deterioro (López-Bote et al., 2003). Los antioxidantes son sustancias que se incluyen en el pienso de los cerdos a baja concentración que retrasan o previenen el deterioro y la aparición de radicales libres de la carne (Thomas et al., 1994), y un tratamiento extra no tiene ningún efecto ni sobre los parámetros zootécnicos ni de rendimiento de canal (Corino et al., 1999; NRC 2012). Pueden actuar de forma individual, vitamina E (acetato de alfatocoferilo) o selenio (Corino et al., 1999; López-Bote et al., 2003) o de manera combina da de 2 o más antioxidantes juntos con acción sinérgica, es decir, evitan la oxidación con mayor efecto que su suma por separado (Duthie et al., 2001). Por lo tanto, el objetivo de este estudio fue investigar, por un lado, el efecto del tipo genético del verraco finalizador y el sexo sobre los rendimientos de crecimiento, canal y contenido en grasa intramuscular, y por otro, el efecto de un suplemento de antioxidantes en el pienso de acabado sobre parámetros de oxidación de la carne y AG subcutáneos de una producción de cerdo ibérico (50 % Duroc) en categoría de “Cebo de Campo”.
MATERIALES Y MÉTODOS
El estudio se realizó en condiciones de cam po, entre diciembre de 2017 y diciembre de 2019, en la unidad de crecimiento-cebo de una granja comercial situada a una Latitud de 38°35’20,3’’ Norte y 7°01’7,8’’ Oeste, en la región de Extremadura (España). Esta zona presenta un clima mediterráneo continentalizado (con cierta influencia atlántica), que se caracteriza generalmente por una importante amplitud térmica, siendo veranos cálidos y secos e inviernos parcialmente fríos y húmedos. Los métodos de manejo de los animales durante la investigación fueron desarrollados de acuerdo con los principios éticos en investigación y docencia con modelos animales aprobados en la Directiva Europea 2010/63/UE publicada en el DOUE (2010) y en las normas nacionales recogidas en el Real Decreto 53/2013 publicado en el BOE (2013), por el que se establecen las normas básicas aplicables para la protección de los animales utilizados en experimentación y otros fines científicos, incluyendo la docencia.
Animales
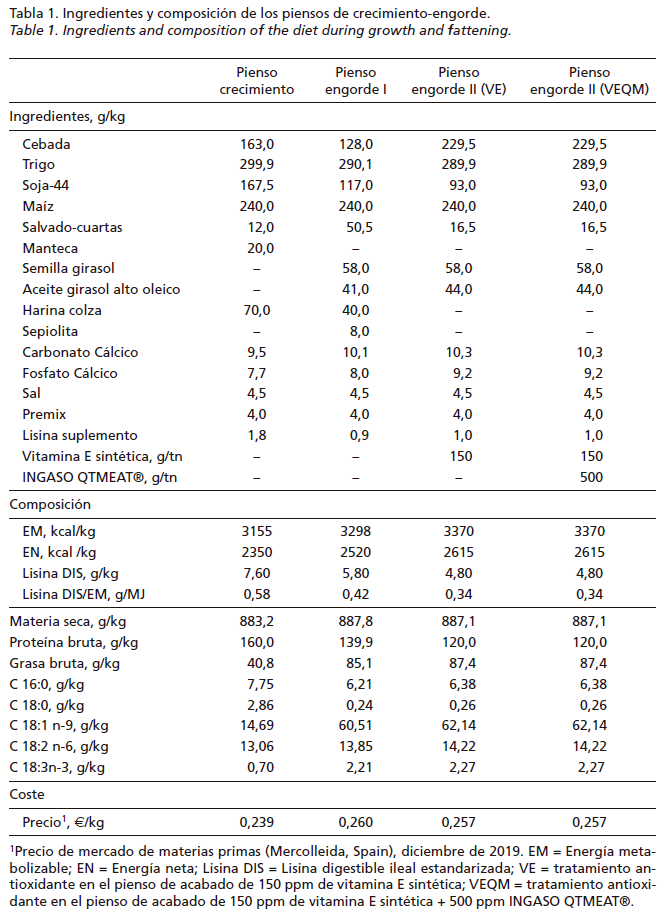
Se utilizaron 2 lotes de lechones ibéricos (50% Duroc) seleccionados al azar a los 98 días de edad en el momento de salida de la transición, nacidos al final de año de 2 años consecutivos en una granja de producción comercial. El lote A constaba de 91 machos castrados de 29,1kg de PV y 95 hembras de 29,3kg de PV, y el lote B de 56 machos castrados de 35,6kg de PV y 60 hembras de 34,3kg de PV. Para la fase de crecimiento, en la misma época del año, ambos lotes se trasladaron a la misma explotación en semi-extensivo donde se evaluaron sus parámetros de crecimiento (Figura 1). Se repartieron en 2 corrales distintos por tamaños homogéneos (machos y hembras mezclados), con las mismas condiciones ambientales, de alimentación y manejo. Los corrales disponían de un área al aire libre suficientemente amplia para caminar con toda libertad, más de 100m2 por cerdo y suelo de tierra. Estos corrales estaban cercados con valla metálica y pared de piedra con una zona cubierta con espacio disponible de 1m2 por cerdo de refugio y descanso. Todos los animales tuvieron libre acceso al agua y restringido al pienso hasta los 303 días de edad y posteriormente ad libitum hasta el sacrificio, 355 días de edad y 170kg de PV. Al sacrificio se seleccionaron, por un lado, 55 hembras y 63 machos del lote A, y 44 hembras y 46 machos del lote B para evaluar parámetros de canal; y por otro, 23 hembras y 36 machos del lote A, y 29 hembras y 32 machos del lote B para analizar parámetros de calidad de carne. Al lote A se asignaron lechones de tipo genético descendientes de DT y fueron alimentados con el pienso de engorde II (VE); al lote B se asignaron lechones de tipo genético descendientes de DTGIM y fueron alimentados con pienso de engorde II (VEQM). La característica diferencial del tipo de verraco Duroc utilizado fue su contenido de GIM medida por ultrasonidos (% GIM_US) en vivo: DT con un 4,26% (población fundacional Duroc Templario) y DTGIM con un 4,49% GIM_US (1ª generación de verracos Duroc Templario). INGASO QTMEAT, es un suplemento nutricional que combina de forma sinérgica diferentes antioxidantes naturales (ácido ascórbico, vitamina E natural, polifenoles naturales y selenio orgánico).
Programa de alimentación
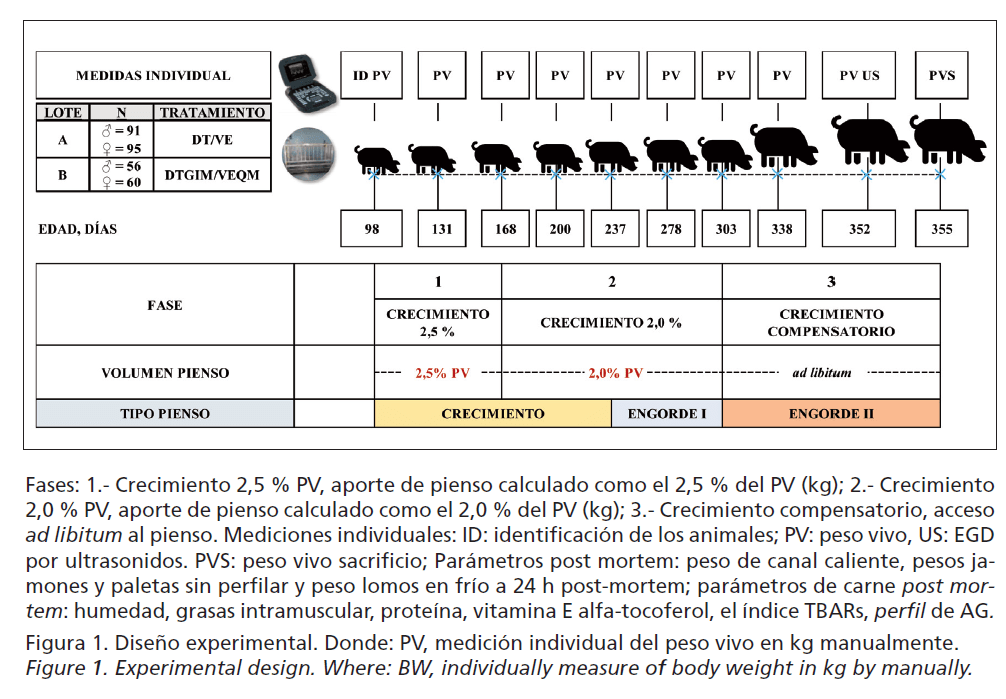
Para la fase experimental de ambos lotes se diseñó un mismo programa de alimentación con el objetivo de obtener una tasa de crecimiento lento global de menos de 550g/d entre los 100 y los 355 días de edad (Figura 1). Los piensos se formularon siguiendo los requerimientos recomendados por FEDNA (2013) y su composición en materias primas y los principales contenidos de nutrientes estimados se muestran en la Tabla 1. Brevemente, los animales siguieron 3 fases de alimentación, crecimiento (98-237 días de edad), engorde I (237-303 días de edad) y engorde II (303-352 días de edad). Siguiendo una práctica habitual en el ecosistema de la dehesa, la cantidad de pienso suministrado se realizó en función de la edad y PV y definirá las 3 fases de crecimiento. Se programó de la siguiente forma: de los 98 a 168 días y de 168 a 237 días de edad “Pienso de Crecimiento” racionado al 2,5 y 2% del PV, respectivamente; de los 237 a 303 días de edad el “Pienso de Engorde I” racionado al 2% del PV; y, por último, de los 303 a 352 días de edad “Pienso de Engorde II” ad libitum (Figura 1).
Parámetros crecimiento y canal
En la Figura 1 aparece el esquema del protocolo de medición de parámetros de crecimiento y canal de los cerdos. Se monitorizaron a lo largo del tiempo mediciones repetidas individuales de PV (kg) para el calculó la ganancia media diaria (GMD; kg/d). Al sacrificio se midió de forma individual el PV y el espesor graso dorsal (EGD; mm) mediante una sonda lineal de ultrasonidos (IMV Imaging) en la posición P2 (última costilla, a 4-6cm de la línea media dorsal) (McCann y Magowan, 2004); en post mortem, el peso de canal en caliente (kg) y en el despiece en frío; y 24h post-mortem, los pesos de piezas nobles: jamones sin perfilar (kg), paletas sin perfilar (kg) y lomos (kg). El rendimiento de canal se calculó en caliente y el de las partes nobles frente a la canal en frío que fue un 2% menos del peso de canal caliente por las pérdidas por oreo durante las primeras 24h de enfriamiento.
Parámetros de calidad de carne
Los parámetros de calidad de carne se midieron en muestras de 20cm de carne recogidas de la zona craneal del músculo longissimus thoracis et lumborum y de 100g de grasa subcutánea recogidas a la altura de la cadera. En la muestra de carne se midió el contenido de: humedad, mediante secado en estufa a 105°C durante 24h o hasta obtener peso constante (Norma ISO 1442:1997); de grasa, usando éter de petróleo (40-60°C) como solvente en una unidad extractora Soxhlet (Soxhlet System HT2 1045 Tecator); y proteína, N Dumas × 6,25 (midiendo el N con un equipo LECO Serie 828) de acuerdo con los AOAC (2006). La determinación del contenido en Vitamina E de las muestras se llevó a cabo según el método descrito por Rey et al. (1996) e Isabel et al.(1999a). La solución tampón utilizada para la extracción estuvo formada por 13,4g de fosfato sódico y 1,86g de EDTA disueltos en 1L de agua y ajustado a pH 7 con HCl 5N. El etanol, metanol y hexano grado HPLC se obtuvieron de PANREAC. Para el análisis cromatográfico se inyectaron 25µl de muestra en un cromatógrafo de líquidos de alta resolución (HPLC) Hewlett-Packard, Serie 1050, equipado con una bomba isocrática y una columna Lichrcart PR 18 (250mm × 4mm y 5µm de tamaño de partícula). La fase móvil empleada fue metanol: agua (97:3) a un flujo de 2ml/min. La detección del alfa-tocoferol se realizó mediante un detector ultravioleta fijado a 292nm. La identificación se realizó por comparación de los tiempos de retención con el correspondiente patrón. La cuantificación se realizó a partir de una cantidad conocida de los distintos patrones y se expresó en forma de µg/g de alimento; de vitamina E alfa-tocoferol, en ppm sobre materia seca (MS) con el equipo de cromatografía líquida HPLC Agilent 1100. La reacción de ácido 2-tiobarbitúrico (TBARS) de las muestras se cuantificó mediante espectrofotometría a una absorbancia de 532nm en una dilución descrita por Botsoglou et al. (1994) a través de una recta de calibrado construida frente a un blanco preparado con la dilución de trabajo de malondialdehído (MDA)(2,39µl/ml) diluida con 2,5ml de una dilución acuosa de ácido tricloroacético (TCA) (5%) + 1,5ml de ácido 2-tiobarbitúrico (TBA) (0,8%). Los análisis de AG se llevaron a cabo siguiendo la metodología expuesta por Isabel et al. (2014), basada en la extracción y metilación de los ésteres metílicos y su posterior inyección y determinación por cromatografía de gases. Los AG fueron fraccionados e identificados utilizando un cromatógrafo Hewlett Packard 6890 con un columna 30mm × 0,32mm × 0,25µm cross-linked polyethylene glycol capillary columna. El programa de temperaturas presentó un rango de 170 a 245°C. El inyector y el detector mantuvieron una temperatura de 250°C. El flujo del gas (helio-portador) fue de 2ml/min. Los AG determinados en el tejido subcutáneo están expresados en porcentaje. En el caso de las dietas están expresados en g/kg a partir de la matriz de formulación. Todos los patrones de AG utilizados fueron suministrados por SIGMA (Madrid) (López-Bote et al., 2003).
Análisis estadísticos
Se realizó un ANCOVA donde los parámetros de crecimiento y de canal observados fueron las variables respuesta (Y), y el sexo (S) y el tipo de verraco terminal (V), las variables explicativas. La estructura del modelo fue la siguiente:
Yijk = µ+ (S)i + (V)j + P0 + εijk, donde:
- Y=datos observados para el cerdo k del factor i j.
- µ = media general.
- S = macho castrado y hembra.
- V = DT y DTGIM.
- P0 = covariable peso inicial: PV1, PV a los 98 días de edad para parámetros de crecimiento (PVi y GMDi); PVS, PV al sacrificio para parámetros de rendimiento de canal y piezas nobles (jamón, paleta, lomo y EGDs).
- ε = error residual.
Además, por un lado, se realizó un ANOVA donde el contenido de grasa intramuscular fue la variable dependientes (Y), y el sexo (S) y el tipo de verraco terminal (V), las variables explicativas. La estructura del modelo fue la siguiente:
Yijk = µ + (S)i + (V)j + εijk, donde:
- Y = datos observados para el cerdo k del factor i j.
- µ = media general.
- S = macho castrado y hembra.
- V = DT y DTGIM.
Y por otro, un ANOVA donde el índice de oxidación lipídica y contenido de AG fueron las variables dependientes (Y), y el sexo (S) y el suplemento antioxidante en el pienso de engorde II (OX), las variables explicativas. La estructura del modelo fue la siguiente:
Yijk = µ + (S)i + (OX)j + εijk, donde:
- Y = datos observados para el cerdo k del factor i j.
- µ = media general.
- S = macho castrado y hembra.
- OX = VE y VEQM.
- ε =error residual.
Los análisis estadísticos se realizaron mediante el software de estadística para Excel (Microsoft), XLSTAT versión 2016. 02.28451 (Addinsoft). Los resultados se presentan como medias y error estándar de la media (SEM) del conjunto de datos en cada caso. El nivel de significación estadística para todos los análisis fue predeterminado (p<0,05).
RESULTADOS
Parámetros crecimiento y canal
La Tabla 2 presenta los resultados del promedio de PV y GMD observados a lo largo del crecimiento y el efecto que tuvo el tipo de verracos terminal y el sexo sobre estas 2 variables. Cabe destacar que el PV inicial (PV1) utilizado como factor covariable en el análisis afectó de manera significativa (p<0,05) en el resultado de PV de cada control y en la tasa de crecimiento global de todo el ciclo. Se observaron diferencias significativas en los PV de cada control (p<0,05) por efecto del tipo de verracos terminal excepto a los 168 días de edad. Sin embargo, no por el sexo hasta los 237 días de edad, momento en el que los machos fueron significativamente (p<0,05) más pesados que las hembras hasta el final del ciclo.
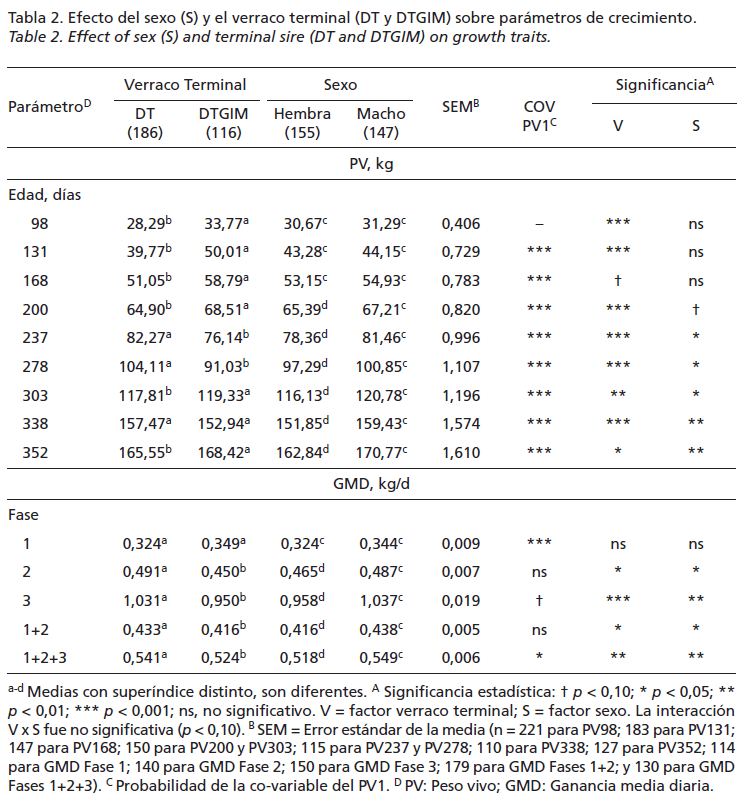
Para la GMD se observaron diferencias signifcativas (p<0,05) tanto por efecto del tipo de verraco como por el sexo en la fase global del ciclo (Fase 1+2+3), la fase de restricción alimentaria (Fase 1+2) de crecimiento lento y la de consumo ad libitum (Fase 3) donde se expresó el crecimiento compensatorio en todos los casos. Los animales descendientes del verraco terminal DT y los machos obtuvieron mayores tasas de GMD que los descendientes del verraco ternial DTGIM y las hembras, respectivamente.
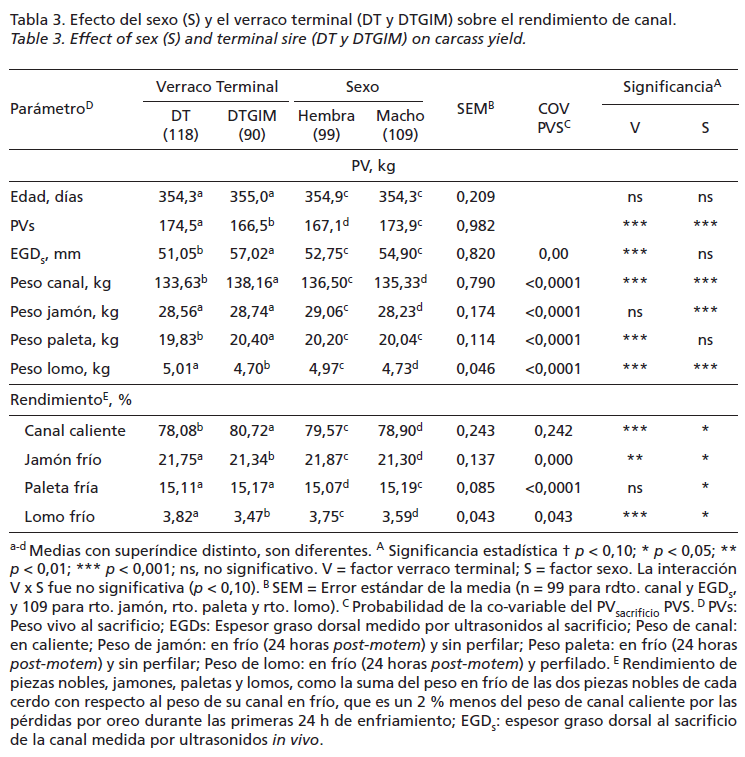
La Tabla 3 recoge los resultados medios de los parámetros de canal medidos, observados para cada factor en estudio (verraco terminal y sexo), observándose que los machos y los animales descendientes del verraco DTGIM obtuvieron valores significativamente (p<0,05) mayores para el EGD, medido por ultrasonidos, y menores para rendimiento de jamón y lomo que las hembras y los animales descendientes del verraco DT, respectivamente. No se observó interacción significativa (p<0,05) entre el tipo de verraco terminal y el sexo y si que el PV sacrificio, utilizado como covariable en el ANCOVA, afecto de manera significativa (p<0,05) sobre los resultados de los 4 parámetros (EGD, jamón, paleta, y lomo) de canal medidos.
Parámetros de calidad de carne
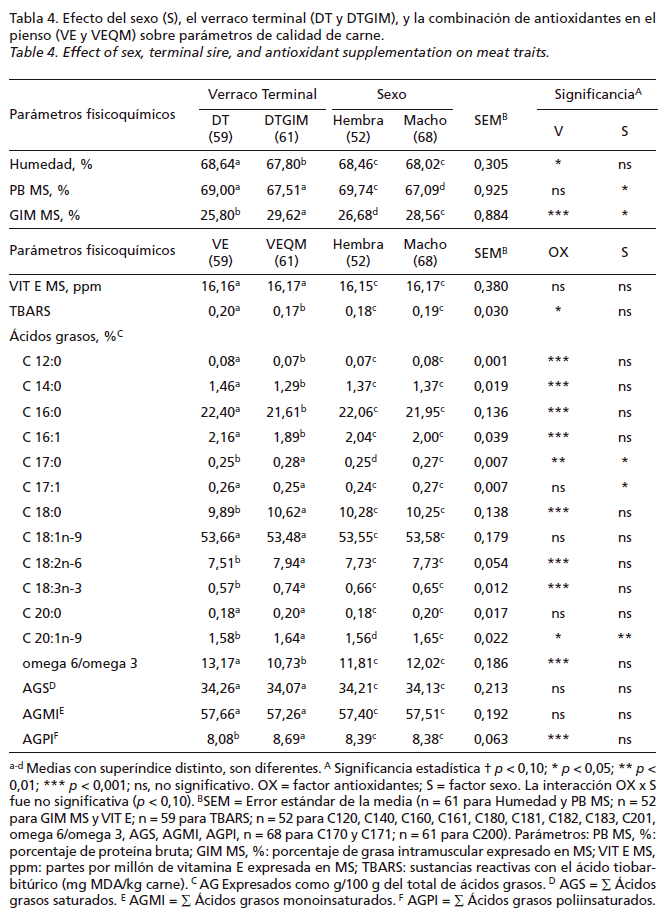
Los resultados y diferencias estadísticamente significativas que se observan por el efecto del verraco terminal y el sexo sobre el contenido de GIM y la combinación de antioxidantes en el pienso sobre el índice TBARS de la carne y la composición de AG de la grasa subcutánea aparecen en la Tabla 4. No se observó interacción entre el tipo de verraco terminal y el sexo para el contenido de GIM. El % Humedad está correlacionado positivamente con el contenido de grasa intramuscular de la carne, de esta manera, las muestras de nuestro estudio presentaron un elevado valor tanto de % Humedad como graso. El efecto del verraco terminal afectó significativamente (p<0,05) sobre el % Humedad, y no el del sexo; se observaron valores mayores en las muestras de animales que descendían del verraco terminal DT (68,64%) que en las de DTGIM (67,80%). Para el contenido de GIM_MS observamos diferencias significativas (p<0,05) tanto por efecto del verraco terminal como por el sexo, los animales que descendían de DTGIM (29,62%) y los machos (28,56%) alcanzaron valores mayores que los DT (25,80%) y las hembras (26,68%), respectivamente. Mientras que para el contenido de proteína bruta de la materia seca (PB MS; %) no hubo diferencias significativas (p>0,05) por efecto del verraco terminal, si las hubo (p<0,05) por efecto del sexo; las hembras presentaron valores mayores de proteína que los machos, 69,74% y 67,09%, respectivamente. En los resultados de índice de oxidación de las grasas (TBARS) se observaron diferencias significativas (p<0,05) por efecto de los antioxidantes en los piensos de engorde II, los VE obtuvieron valores significativamente (p<0,05) mayores que los de VEQM (0,20 y 0,17; respectivamente). En cuanto a los valores de vitamina E alfa-tocoferol determinada en músculo longissimus thoracis et lumborum sobre la MS, no se encontraron diferencias significativas (p>0,05) por efecto de ningún factor estudiado. Para la composición de AG de la grasa subcutánea, no se observaron diferencias significativas (p>0,05) en el contenido de AG saturados (AGS) y AG monoinsaturados (AGMI) para ningún factor estudiado. Y en el contenido de los AG polinsaturados (AGPI) y ratio AG omega-6:omega-3 sí hubo diferencias significativas (p<0,05), los animales VEQM presentaron mayores valores de AGPI que los VE (8,69% versus 8,08%) y menores de ratio omega-6:omega-3 (10,73% versus 13,17%, respectivamente).
DISCUSIÓN
Parámetros de crecimiento y canal
En el actual estudio, el ritmo de crecimiento de la fase de restricción alimentaria (Fase 1+2) desde los 98 hasta los 303 días de edad dependió de la cantidad de pienso aportado y el sexo a partir de los 168 días, momento en el que se reduce el aporte de pienso y aumenta la competencia por el mismo, en estas circunstancias los machos fueron favorecidos sobre las hembras. A su vez, en la fase de realimentación ad libitum (Fase 3) la GMD fue afectada significativamente por el tipo de verraco terminal y el sexo, no por el peso de inicial. En esta última fase del ciclo, los animales descendientes de verraco DT mostraron un significativo (p<0,05) mayor potencial de crecimiento (1,031 vs. 0,950kg/día) que los de DTGIM. Y los machos alcanzaron tasas de crecimiento (1,037 vs. 0,958 kg/día) significativamente mayores que las hembras, a diferencia con los resultados obtenidos por otros autores como Usero et al. (2016) y Serrano et al. (2008), que no encontraron diferencias significativas ni por efecto del genotipo del verraco finalizador ni por el sexo en la fase de crecimiento entre los 19-145kg de PV del cerdo ibérico (50% Duroc) de “Cebo” criado en condiciones de intensivo. Por un lado, los resultados de ritmo de crecimiento lento medios obtenidos en la fase de restricción alimentaria coinciden con Daza et al. (2008), que indica que la cantidad de la reducción del crecimiento provocada por la restricción alimentaria es proporcional al nivel de restricción excepto cuando los animales están expuestos a temperaturas medias por debajo la termoneutra, como de hecho sucedió en la Fase 2, que transcurrió durante el invierno, donde la velocidad de crecimiento se redujo; por otro, los resultados de GMD obtenidos en la fase de realimentación ad libitum de crecimiento compensatorio coinciden con Campbell y Dunkin (1983), que encontraron que los cerdos con restricciones tienen un mayor rendimiento digestivo que los alimentados ad libitum como consecuencia de una reducción de los gastos de mantenimiento, lo que explica en parte el efecto de su mayor eficiencia en el uso del pienso. Este efecto se prolonga durante 2-3 semanas si el periodo de restricción va seguido de una alimentación ad libitum, provocando un mayor consumo y ritmo de crecimiento, que se prolonga durante 6 semanas, momento que alcanzan los 160kg de PV de sacrificio. En estudios previos, se observó que el efecto del crecimiento compensatorio puede aparecer de forma más marcada por efecto del “fenotipo ahorrador”, característico de la raza ibérica y se describe como una estrategia diseñada para aumentar el crecimiento de peso y la deposición de grasa tras una restricción energética (Daza et al., 2008). Los animales descendientes del verraco terminal DTGIM, significativamente más grasos que los DT (57,02 vs. 51,05mm EGD), tuvieron un significativo mayor rendimiento de canal (80,72 vs. 78,08%) y paletas (15,17 vs. 15,11%), de acuerdo con los resultados encontrados por Serrano et al. (2008), y menores de rendimiento de jamón (21,34 vs. 21,75%) y lomo (3,47 vs. 3,82%), esta vez en descuerdo con este trabajo de Serrano. En cuanto al sexo, los machos obtuvieron valores de EGD de la canal significativamente (p<0,05) mayores que las hembras (54,90 vs. 52,75mm EGD) y menores de rendimiento de jamón (21,30 vs. 21,87%) y lomo (3,59 vs. 3,75%), de acuerdo con Serrano et al. (2008). En general, las canales menos grasas obtuvieron mejores rendimientos de jamón y lomo, y peores de paleta, discrepancias que podrían estar relacionadas con la diferente metodología utilizada por otros autores para el despiece llevado a cabo de las canales.
Contenido de grasa intramuscular
Los resultados medios de contenido de GIM_MS (%) obtenidos en este trabajo están de acuerdo con los obtenidos por Vázquez-Gómez et al. (2020) y son mayores que los publicados por Barbero et al. (2013) en cerdo ibérico (50% Duroc) de “Cebo”, y están de acuerdo con Serrano et al. (2008) en su estudio en cerdo ibérico (50% Duroc) de “Cebo” en intensivo, donde se observó que hubo efecto del tipo de verraco y el sexo sobre su contenido. En nuestro estudio, los machos obtuvieron mayores valores que las hembras, de acuerdo con Egea et al. (2016) y Serrano et al. (2008), y en discrepancia con los resultados obtenidos por Usero et al. (2016) en cerdo ibérico (50% Duroc) de “Cebo”.
Oxidación lipídica y composición de los ácidos grasos
La oxidación lipídica es la principal causa del deterioro de la carne. Sobre los resultados del índice de oxidación de las grasas (TBARS) se observó el efecto del nivel de antioxidantes en el pienso. De cualquier modo, para ambos lotes experimentales, estaban lejos de valores de 0,5; considerado valor crítico para la estabilidad oxidativa de la carne (Wood et al., 2008). Los valores obtenidos de vitamina E resultaron de acuerdo con los obtenidos por Isabel et al. (1999a), de 16,87 ppm en MS, en músculo biceps femoris de cerdo blanco suplementado con 200ppm de vitamina E sintética en el pienso durante los últimos 42 días de vida. Aumentar la vitamina E en la dieta es una estrategia para aumentar la estabilidad oxidativa y mejorar las características sensoriales de los jamones curados (Isabel et al., 1999b), pero aunque la vitamina E tiene muy poca toxicidad, por mucho que se aumente su nivel de inclusión en el pienso, esta alcanza un punto que no responde a una mayor acumulación en el músculo (Bonnette et al., 1990). Así, en este estudio se observó que no había diferencias significativas en el contenido de vitamina E muscular postmortem en animales VEQM (150ppm vitamina E sintética + 18,2ppm de vitamina E natural extra) con respecto a los VE (150ppm vitamina E sintética). Por eso, más allá de aumentar la vitamina E para mejorar la protección antioxidante de la carne, el programa de alimentación experimental siguió la estrategia de estudiar combinaciones sinérgicas de otras moléculas naturales con características antioxidantes junto con la vitamina E. Además, se diseñó para dirigir el perfil de AG a controlar la síntesis endógena de AGS y estimular la acumulación de AGMI y AGPI, más sensibles a la oxidación. Para ello, se formularon los piensos de crecimiento y engorde con niveles relativamente bajos de proteína y lisina, y altos de energía, grasa, AGMI y AGPI, en comparación con piensos convencionales (Estévez et al., 2006). Y se llevó a cabo un racionamiento dirigido por fases explicado anteriormente en la Figura 1 (Daza et al., 2007a). En este sentido, se obtuvieron valores bajos de C16:0 y C18:0 por debajo del 22,5% y 11%, respectivamente, y altos de C18:1, por encima del 53%, similares a los estudiados (Ninoles et al., 2007; Daza et al., 2007b) en cerdo ibérico (100 %) de “montanera” y diferentes de los observados por Serrano et al. (2008) en su estudio realizado en cerdo ibérico (50% Duroc) “Cebo”. En este caso, eran cerdos alimentados con piensos con una composición diferente para el crecimiento: (19 a 80kg PV), concentración energética de 9,15MJ/kg, lisina total del 0,72% y grasa mezcla animal del 0,8%; el cebo (80 a 110kg PV): concentración energética de 9,74MJ/kg, lisina total del 0,64 %, jabón cálcico en base a oleínas del 1% y grasa mezcla animal del 2,10%; y el acabado (110 a 145kg PV): concentración energética de 10,38MJ/kg, lisina total del 0,51%, jabón cálcico en base a oleínas del 3,00 % y grasa mezcla animal del 3%. Sin embargo, los valores de C 18:2 obtenidos en el presente experimento alcanzan el mismo nivel que en las investigaciones de Ninoles et al. (2007) y Serrano et al. (2008), y menores que los obtenidos por Daza et al. (2007b); este suceso se explica por la diferencia de consumo de este AG en cada caso, ya que la concentración de C18:2 en la grasa del cerdo es directamente proporcional a la cantidad consumida (Wood et al., 2008; Isabel et al., 2014). Este AG juega un papel esencial en la consistencia de la grasa y su menor acumulación en cerdo ibérico está relacionada tanto por su consumo como por su capacidad de utilizarlo a través de la beta-oxidación (Isabel et al., 2014). Por otro lado, por el factor de los antioxidantes no se encontraron diferencias significativas en el pienso sobre el contenido de los AG de síntesis endógena y de resultado de la desaturación, AGS y AGMI, respectivamente (Óvilo et al., 2014), y sí sobre el contenido de AGPI absorbidos por la dieta. Los animales VEQM con respecto a los VE obtuvieron un contenido significativamente mayor de todos los AGPI analizados AG C 18:2, C 18:3 y una relación omega-6:omega-3 significativamente menor por el destacable efecto antioxidante sobre estos AG especialmente sensibles a la oxidación (Cholewski et al. 2018), destacando el contenido de omega-3 de un 0,74% en VEQM respecto al 0,57 % en los VE, por lo que la suplementación en la dieta con antioxidantes adicionales son necesarios para su mayor absorción (Heras-Molina et al., 2020). En este sentido, la estrategia de la administración de polifenoles de la uva junto a la vitamina E pudo mejorar la disponibilidad de estos AG de la dieta, del mismo modo que el hidroxitirosol disminuye la peroxidación de los lípidos e incrementa la disponibilidad fetal de AG omega-3 y omega-6 poliinsaturados (García-Contreras et al., 2019). Por lo tanto, la estrategia de alimentación seguida en este estudio fue de especial interés, encontrando rutas para manipular la composición de AG de la carne en virtud de producir alimentos funcionales (Coates et al., 2009). Dietas ricas en ácido oleico y una baja proporción omega-6:omega-3 muestran propiedades antiinflamatorias (Fernández et al., 2018), reducen los niveles de colesterol y se asocian con una menor incidencia en enfermedades cardiovasculares (Mattson y Grundy, 1985). En general, bajar la relación omega- 6:omega-3 en la dieta a 5:1 puede ser beneficioso para los perfiles lipídicos en la sangre, mejorando el valor nutricional de la carne de cerdo, sin efecto adverso sobre el rendimiento del crecimiento y los parámetros de calidad de la carne (Liu y Kim, 2018).
CONCLUSIONES
El presente estudio sugirió que el tipo genético obtenido aplicando selección genética mejoró la cantidad y calidad de la grasa. Además, que la utilización de una combinación de antioxidantes en el pienso pareció efectiva para incrementar el valor nutricional de la carne y protegerla frente a la oxidación. Estos hallazgos sirven para aumentar el valor añadido a la carne de cerdo ibérico en su faceta “Cebo de Campo”, otorgándole propiedades nutricionales y sensoriales de gran interés
para el consumidor.
Autores
-
Guillermo Ramis-Vidal, Antonio Muñoz-Luna. Departamento de Producción animal, Facultad de Veterinaria de la Universidad de Murcia, España.
-
Guillermo Usero-Alonso, Francisco Gil-Rueda y Antonio Muñoz-Luna. FARM Faes Nutrición y Salud Animal, 01308 (España).
Deja un comentario