

La producción de por vida y la longevidad son parámetros muy importantes de rentabilidad en la cría de las reproductoras. La oportunidad de mejorar el rendimiento y la longevidad de por vida se puede encontrar en el período de cría y en la preparación de las primerizas para su futura función reproductora. Con el objetivo de prevenir el desecho prematuro, es posible influir en la condición corporal, la condición de las patas, el desarrollo de la glándula mamaria, y el funcionamiento adecuado del tracto reproductivo a través de estrategias de nutrición, tecnología, y crianza. La nutrición juega un papel muy importante, ya que puede afectar a todos los requisitos básicos para lograr un desempeño satisfactorio de las primerizas.
La selección de la estrategia de cría más eficaz puede resultar difícil porque hay muchos factores que afectan la producción y la longevidad. El objetivo de esta revisión bibliográfica es proporcionar información actualizada sobre cómo se puede influir en la longevidad y la producción de las cerdas mediante la elección de estrategias de cría de las primerizas, y la importante área de la nutrición.
INTRODUCCIÓN
Además del tamaño y el peso de la camada, la longevidad es un indicador crucial de la rentabilidad del rebaño de cerdas. Por lo tanto, es importante crear condiciones óptimas para las cerdas en las fases individuales de sus ciclos reproductivos. Incluso si los productores pueden elegir nulíparas en condiciones físicas óptimas, con un número suficiente de pezones y ubicarlas en un ambiente casi óptimo, esto aún no es garantía de lograr el éxito reproductivo y la longevidad. Es importante comenzar a prestar especial atención a las primerizas mucho antes, ya que se están criando antes de incluirlas en el rebaño de reproducción para garantizar un desarrollo corporal adecuado y el inicio de las funciones reproductivas.
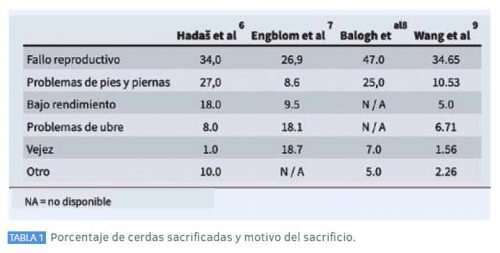
La longevidad está asociada con el nivel de cerdas sacrificadas. Aunque el reemplazo anual del 40% de las cerdas se considera económicamente recomendable, varía dentro de un amplio rango (62% para algunas granjas de EE.UU. en 2019) y depende de las condiciones y el manejo de cada rebaño. Incluso los niveles de reemplazo anual más altos pueden ser económicamente aceptables si las hembras del rebaño reproductor son lo suficientemente productivas; sin embargo, el bienestar animal y la viabilidad económica a largo plazo pueden ser motivo de preocupación cuando los niveles de reemplazo están por encima del 50%. En un resumen de 2018 para los EE.UU., informaron de una tasa media de eliminación selectiva del 45,06%. La tasa de sacrificio total incluyó sacrificio voluntario e involuntario. Para el sacrificio voluntario, Mote et al, recomendó obtener al menos 3 camadas de cada cerda para recuperar la inversión en la cerda. La selección de cerdas que puedan permanecer en el hato reproductor durante más tiempo es beneficiosa para el rendimiento reproductivo. Los autores asumen que las principales razones para el sacrificio no cambian sustancialmente con el tiempo, y esto ha sido documentado por publicaciones a lo largo de los años. Friendship et al, mencionaron los trastornos reproductivos (43%), los problemas de las extremidades (12%) y el bajo rendimiento (7%) entre las causas más frecuentes de sacrificio. Stupka et al, informaron que las causas más frecuentes de sacrificio de las explotaciones porcinas son problemas reproductivos (44%), problemas musculoesqueléticos (19%) y otras razones como la producción de leche, el estado de salud y la edad (28%). Hadaš et al, realizó una evaluación según el orden de parto y encontró que los niveles más altos de descarte se alcanzaron después del 1º y 2º parto, con 22% o 21% de cerdas descartadas del hato de cerdas, respectivamente, con fallas reproductivas (34%), trastornos musculoesqueléticos (27%), y bajo rendimiento (18%) siendo las causas más frecuentes de descarte.
El mal estado de las glándulas mamarias y el estado de salud representaron cada uno menos del 10% de los casos. El porcentaje de cerdas sacrificadas y el motivo del sacrificio se enumeran en la Tabla 1. Estos motivos para el sacrificio indican las áreas que presentan margen de mejora durante la crianza y preparación de las primerizas. Sin embargo, un alto nivel de sacrificio involuntario también puede ser un indicador de poca habilidad del personal o deficiente bienestar de las cerdas.
La selección para mejorar la prolificidad ha dado como resultado camadas más grandes y, por lo tanto, aumentó la proporción de lechones con bajo peso al nacer (BPN). Está documentado que los lechones de bajo peso al nacer tienen un rendimiento de crecimiento-finalización y una calidad de la canal más pobres. El peso al nacer también tiene una relación con el desempeño reproductivo posterior en primerizas. Almeida et al, investigó los efectos del peso al nacer en el tracto reproductivo y el desarrollo del folículo ovárico en primerizas de 150 días de edad. Se criaron hasta los 150 días de edad 28 cerdas de diferentes rangos de peso al nacer (alto peso al nacer [APN]: 1,8-2,2kg; bajo peso al nacer [BPN]: 0,8-1,2kg) de cerdas comerciales de mayor número de partos. Sus pesos corporales (PC) se registraron al destete, al final de la transición y al final de las fases de crecimiento y finalización. Las primerizas con BPN mostraron un peso corporal significativamente más bajo y una ganancia diaria promedio más lenta durante todas las fases de producción en comparación con las del grupo APN (P<0,01). La mayoría de las medidas biométricas del tracto reproductivo fueron similares entre los grupos experimentales, excepto que la longitud vaginal y el índice gonadosomático (peso ovárico relativo) se vieron afectados por la clase de peso al nacer (P< .05). Las hembras con bajo peso al nacer también mostraron menos folículos ováricos de tamaño mediano (3-5 mm; P<0,01), tendieron a tener menos folículos preantrales (P<0,07) y más folículos atrésicos por área de la corteza ovárica (P<0,05). Por lo tanto, además de afectar el rendimiento del crecimiento posnatal, el peso al nacer influyó en la longitud vaginal y la dinámica folicular, lo que puede afectar al rendimiento reproductivo de las primerizas de reemplazo.
De manera similar, Vallet et al encontraron que la longitud uterina total se asoció positivamente con los pesos al nacer. Sus resultados indican que el consumo de calostro, el peso al nacer, la tasa de crecimiento antes del destete, el número de destetados y la paridad se asociaron con los rasgos de desarrollo de las primerizas durante la vida posterior. Knauer, encontró que el mayor peso al nacer de los lechones estaba relacionado con la proporción de primerizas que parían una camada. El mayor crecimiento de los lechones antes del destete se relacionó con la proporción de primerizas que parieron una camada y el rendimiento reproductivo de por vida. Por lo tanto, las estrategias de manejo que mejoran la producción de calostro, la producción de leche y el crecimiento de lechones antes del destete deberían mejorar la productividad de por vida posterior. El aumento de la edad de destete en un día se sumó a la reproducción posterior de una primeriza en 0,185 lechones/año, y las primerizas que recibieron crianza cruzada tuvieron un 2,45% menos de probabilidades de parir una camada.
NUTRICIÓN MINERAL
Es bien entendido que la nutrición juega un papel integral en el desarrollo de una primeriza. Las primerizas deben criarse en lugar de cebarse, por lo que las dietas diseñadas para cerdos de finalización pueden no satisfacer las necesidades fisiológicas de las primerizas de reemplazo. Las primerizas de reemplazo en la fase de crecimiento-acabado deben recibir dietas diseñadas específicamente. Los genotipos de líneas maternas modernas son más sensibles al manejo nutricional porque su apetito es menor y tienen un potencial de crecimiento magro excepcional. Por lo tanto, las primerizas de hoy son más susceptibles a las deficiencias en nutrición, medio ambiente y manejo.
Para lograr un mejor desempeño en la crianza de las cerdas y un mejor crecimiento de sus cerdos, se requiere un suministro adecuado de minerales, incluidos los oligoelementos. La base y el desarrollo esquelético, el peso al nacer, la producción de leche y el crecimiento pueden verse afectados negativamente cuando los minerales no satisfacen las necesidades del animal. Los requerimientos de las cerdas de calcio (Ca), fósforo (P), sodio y cloro, así como de zinc, yodo y selenio no se satisfacen con alimentos vegetales naturales, por lo que es necesario complementarlos.
Uno de los principales objetivos de la nutrición de nulíparas de reemplazo es aumentar las reservas de minerales maximizando la mineralización ósea. Las dietas para cerdos de finalización pueden no proporcionar el equilibrio correcto de minerales para satisfacer los requisitos nutricionales para el rendimiento reproductivo y para la formación e integridad de cartílagos y huesos. En general, se recomienda proporcionar Ca y P en niveles superiores a los que normalmente se encuentran en las dietas de crecimiento y finalización para evitar que las hembras experimenten problemas de locomoción más adelante debido al agotamiento excesivo de las reservas de minerales durante los períodos de lactancia. Johnston afirma que se ha demostrado que el aumento de la mineralización ósea aumenta la longevidad de las cerdas.
En las dietas de desarrollo de primerizas, se necesita una relación Ca:P digestible mínima de 1:1, y varía según el nivel de P. Por ejemplo, puede ser 1,25:1 si P cumple con las recomendaciones de 50 a 80kg de peso vivo. Además, las recomendaciones de Ca para maximizar la mineralización ósea son mayores que para el crecimiento (menos de 1,35:1 si la concentración de P está en el requerimiento). Aunque las primerizas en crecimiento generalmente tienen acceso ad libitum al alimento, las rápidas tasas de crecimiento en las líneas genéticas actuales y la alta incidencia de problemas en las patas pueden provocar cojera. Los trastornos de cojera representan el 22,5% de los sacrificios de cerdas, y la cojera es una de las causas más importantes de la reducción de la longevidad y el bajo bienestar de las primerizas de reemplazo. El problema se ve agravado por alojamientos y dietas inadecuados durante el período de crianza. Los intentos de mejorar la integridad del esqueleto mediante la reducción de la tasa de crecimiento a través de la restricción de energía no han tenido éxito. Por lo tanto, se requiere la aplicación de herramientas de manejo que sean consistentes con los procesos fisiológicos para reducir los problemas de cojera.
La deficiencia de AP puede hacer que la tasa de crecimiento y la mineralización ósea sean subóptimas, aunque sin efectos sobre la osteocondrosis (OCD). La osteocondrosis es una causa frecuente de cojera y, en consecuencia, una razón para sacrificar cerdas jóvenes. La selección genética podría usarse para reducir la prevalencia del TOC, aunque esto puede ser difícil inicialmente porque el potencial de crecimiento del tejido magro está genéticamente asociado con el TOC. No se conocen bien otros factores que podrían influir en la progresión del TOC. Las estimaciones de heredabilidad de la puntuación del TOC fueron similares para las razas Landrace y Yorkshire, con un promedio de alrededor de 0,21, en un estudio genético realizado por Yazdi et al. Las correlaciones entre los valores genéticos para la longevidad y el TOC fueron bajas (en promedio 0,07, ajustado por las tendencias genéticas) pero significativas (P<0,01) y en una dirección favorable, ya que un mayor TOC se asoció con un mayor riesgo de ser sacrificado.
Fabà et al, complementaron la dieta básica de nulíparas en crecimiento con microminerales orgánicos (cobre, manganeso y zinc a 10, 20 y 50mg/kg, respectivamente) y observaron que esto mejoraba la fortaleza ósea y la densidad ósea. Otra dieta con metionina adicional (a una proporción de metionina: lisina del 102%) aumentó la proporción de hueso muy denso (medido por los valores de Hounsfield). La combinación de estos dos tratamientos dietéticos redujo las puntuaciones de lesiones del TOC en comparación con la dieta basal.
Quinn et al, informaron mejores puntajes de locomoción, mayor densidad mineral ósea y menores puntajes de lesiones de cartílago en primerizas alimentadas con una dieta restringida formulada para grasa en lugar de deposición magra (con mayor contenido de energía y menor contenido de lisina que una dieta de finalización) y con niveles aumentados de cobre, zinc y manganeso. Hartnett et al, utilizó manganeso, zinc y cobre al 206%, 122% y 179%, respectivamente, de las recomendaciones del Consejo Nacional de Investigación para cerdas gestantes y lactantes. Los beneficios de complementar estos minerales podrían conducir a mejoras potenciales en el rendimiento de por vida de las primerizas de reemplazo y la longevidad de las cerdas. Existe una clara indicación de que las primerizas de reemplazo pueden beneficiarse en términos de salud de las extremidades y su bienestar general al ser criadas en corrales exclusivos para hembras (ya que las primerizas criadas con cerdos de finalización machos intactos están expuestas a altos niveles de monta sexual y agresión, lo que puede causar daños físicos) y una dieta suplementada con minerales.
Aunque las deficiencias nutricionales reducen la calidad de los huesos y pueden influir en el TOC, los hallazgos inconsistentes de la investigación en esta área plantean dudas sobre el potencial de los suplementos nutricionales. Estas medidas dietéticas pueden actuar potencialmente para prevenir el TOC o revertir las primeras etapas del TOC, pero no pueden usarse para curar las etapas avanzadas del TOC. Se necesita más investigación para comprender la patogenia y la progresión del TOC, y las interacciones con la tasa de crecimiento, la genética y el manejo.
DESARROLLO DE LA GLÁNDULA MAMARIA
Otro factor importante para un buen desempeño reproductivo y longevidad es la producción suficiente de leche. Las mejoras en la producción de leche de las cerdas a lo largo de los años se han logrado principalmente a través de la nutrición y el manejo porque un estudio reciente demostró que 21 años de selección genética (de 1977 a 1998) aumentaron el peso al nacer de los lechones pero no tuvieron efecto en la producción de leche de las cerdas. Por lo tanto, es necesario diseñar estrategias de manejo que optimicen la producción de leche, y una forma posible es influir en el desarrollo de la glándula mamaria. El número de células mamarias presentes al inicio de la lactancia tiene un gran impacto en la producción potencial de leche de las cerdas33. Varios estudios han demostrado que la nutrición de las nulíparas en los períodos de rápida acumulación mamaria que ocurren durante la prepubertad, la gestación y la lactancia pueden afectar el desarrollo mamario34. Diversos tratamientos nutricionales pueden provocar un aumento del 27 al 52% en el peso del tejido mamario. Un estudio en el que se impuso una restricción alimenticia del 20% en el período prepuberal mostró que la masa del parénquima mamario disminuyó en un 26,3%. La alimentación ad libitum durante el período prepuberal aumentó el peso del parénquima mamario entre un 36 y un 52%. Se estableció claramente que la restricción alimenticia desde los 90 días de edad (pero no antes de los 90 días) hasta la pubertad tenía efectos perjudiciales en el desarrollo mamario de los cerdos.
Según Farmer et al, las primerizas que eran obesas (36mm de grasa dorsal) o demasiado delgadas (12-15mm de grasa dorsal) al final de la gestación tenían tejido mamario menos desarrollado. Las primerizas de peso corporal similar en el momento del apareamiento recibieron diferentes cantidades de alimento a lo largo de la gestación (1,30, 1,58 o 1,82 veces los requisitos de mantenimiento) para lograr 3 niveles de espesor de grasa dorsal (BF) en el día 109 de gestación, a saber, 12 a 15mm (magra), 17 a 19mm (medio), y 21 a 26mm (grasa). La masa de tejido parenquimatoso se redujo significativamente en las nulíparas magras, con 1.059, 1.370 y 1.444g, respectivamente, para las nulíparas magras, medianas y gordas. Estos hallazgos demuestran que, dentro de este rango de condiciones corporales, ser demasiado delgada al final de la gestación es perjudicial para el desarrollo mamario, mientras que las condiciones corporales medianas o grasas no tuvieron un impacto negativo. Se debe evitar la subalimentación para garantizar la cantidad máxima de masa de tejido parenquimatoso. La sobrealimentación energética al final de la gestación también parece ser perjudicial. Se llevó a cabo un experimento para estudiar el efecto de la ingesta de proteínas durante el período de crecimiento y finalización sobre el desarrollo mamario en primerizas. La reducción de la proteína cruda de la dieta del 18,7 al 14,4% desde los 90 días de edad hasta la pubertad no afectó la mamogénesis. No se alteró la cantidad de tejido parenquimatoso ni la composición del parénquima mamario. Esto sugiere que el consumo total de alimento es más importante que el consumo de proteínas para garantizar el desarrollo mamario adecuado de las nulíparas en crecimiento.
Aunque se han realizado investigaciones para evaluar el control nutricional de la mamogénesis en cerdos, es evidente que queda mucho por aprender antes de que se pueda formular la mejor estrategia nutricional para mejorar el desarrollo mamario. Alimentar ciertos extractos de plantas con propiedades estrogénicas o hiperprolactinémicas también puede resultar beneficioso para estimular el desarrollo mamario dentro de períodos fisiológicos específicos. Se hizo un intento de estimular el desarrollo mamario en primerizas proporcionando una fuente dietética de estrógeno. Cuando se añadieron 2,3g/día del fitoestrógeno genisteína a una dieta estándar basada en harina de soja de nulíparas en crecimiento de 90 a 183 días de edad, hubo un aumento del 44% en las células del parénquima mamario al final del período de tratamiento. La genisteína es una isoflavona que se encuentra en las legumbres, especialmente en la soja. En otro estudio, Farmer et al, utilizaron el extracto de la planta silimarina (de Silybum marianum, generalmente conocido como cardo mariano). 4 gramos de silimarina se administraron 2 veces al día a primerizas de 90 a 110 días de gestación, momento en el cual los animales fueron sacrificados para recolectar sus glándulas mamarias. Aunque la alimentación con silimarina condujo a un aumento del 51,8% en las concentraciones de prolactina circulante 4 días después del inicio del tratamiento, este aumento fue transitorio y no fue lo suficientemente grande como para provocar efectos beneficiosos sobre el desarrollo mamario. Las micotoxinas en los alimentos pueden afectar el desarrollo de las glándulas mamarias y del tracto reproductivo muy probablemente a través de sus actividades similares a las del estrógeno. Stephan et al, encontraron que las micotoxinas se transmitían a través de la leche de las cerdas a los lechones sobre la base de la concentración de zearalenona/a-zearalenol en la bilis de los lechones y una tendencia a un menor peso del útero entre los lechones que tenían influencia de zearalenona durante la gestación y la lactancia.
El número de pezones es un criterio importante para las primerizas de reemplazo. Según Drickamer et al, el número de pezones de los cerdos está significativamente influenciado por la genética, principalmente por parte de la madre. La proporción de machos en una camada parece estar relacionada con la distancia anogenital de las primerizas, posiblemente como resultado de un efecto de posición intrauterina. Un mayor número de pezones en la madre y una menor proporción de machos en la camada se asociaron con un mayor número de pezones en la primeriza. Concentraciones de nutrientes y estrategia de alimentación En comparación con las dietas típicas para cerdos en finalización, las dietas para primerizas de reemplazo deben contener concentraciones más altas de vitaminas A y E, calcio, fósforo, selenio, cromo y zinc porque las primerizas altamente prolíficas llegan a la pubertad con reservas limitadas de proteína y grasa corporal y continúan creciendo durante la pubertad y su primera gestación. Una premezcla de vitaminas debe contener niveles elevados de vitaminas liposolubles A, D, E y K, así como vitaminas hidrosolubles colina, biotina y ácido fólico, cuyos niveles son relativamente bajos o están ausentes en las dietas típicas de finalización.
La densidad energética y de aminoácidos de las dietas para cada fase de crecimiento dependerá del potencial de crecimiento magro de las primerizas y del consumo voluntario de alimento. Por lo general, a las primerizas de reemplazo se les proporciona acceso ad libitum a una dieta más baja en energía, proteínas o ambas que las dietas que se suministran a los cerdos de sacrificio para evitar el exceso de grasa corporal. Esto también permite un crecimiento un poco más lento, lo que limita el tamaño del cuerpo maduro y, por lo tanto, previene los problemas de pies y piernas y la acumulación excesiva de grasa. Long et al, informaron que las cerdas alimentadas con una dieta alta en energía y alta en proteínas ad libitum de 120 a 180 días de edad tenían una longevidad significativamente menor a través de 4 partos que las primerizas alimentadas con una dieta alta en energía y baja en proteínas ad libitum o una dieta alta en proteínas con alimentación restringida (35% frente a 56 y 55%, respectivamente). De manera similar, Hoge y Bates encontraron que las primerizas de crecimiento más lento tenían un menor riesgo de ser sacrificadas en su estudio.
Alimentar ad libitum a las primerizas modernas con alto contenido magro es más práctico para la mayoría de los sistemas de producción, especialmente cuando las primerizas se alojan en grupos. Limitar la alimentación puede ser más apropiado para primerizas maternas de bajo y medio peso. Limitar la alimentación implica proporcionar a las primerizas de reemplazo ad libitum acceso a una dieta hasta 1 o 2 meses antes de la reproducción. Las dietas ad libitum son similares a las dietas de crecimiento y finalización, lo que permite la máxima expresión del potencial genético del animal para la tasa de crecimiento y la grasa dorsal. El consumo de alimento se restringe entonces a aproximadamente 85 a 90% de ad libitum, hasta 10 a 14 días antes del apareamiento. Al restringir la dieta, se debe restringir la energía, pero no los aminoácidos, las vitaminas o los minerales. Por lo tanto, las concentraciones de estos nutrientes deben ajustarse al alza en las dietas en consecuencia. El diseño de las instalaciones puede dificultar que los productores alimenten con una dieta restringida a las hembras de reemplazo. Cuando las primerizas se alojan y alimentan en grupos, es difícil asegurar que se ingiere la cantidad correcta de alimento de forma individual porque todas las primerizas no consumen el mismo ritmo. A menos que los productores tengan puestos individuales o un sistema de alimentación electrónico disponible para las hembras potenciales de reemplazo del rebaño reproductor, será difícil implementar un programa de alimentación restringido. Alimentar con una dieta alta en fibra que sea más baja en concentración de energía es una alternativa que permite una ingesta diaria de alimento más cercana a los niveles ad libitum. Los efectos del aumento del tiempo de consumo, el llenado intestinal y la saciedad pueden aliviar parcialmente la competencia y la variabilidad en el consumo de alimento individual en situaciones de alimentación grupal, pero también pueden presentar desafíos relacionados con los sistemas de suministro de alimentos y el manejo del estiércol. El contenido de fibra dietética es significativo debido a la saciedad, la digestión adecuada y el efecto sobre la microflora intestinal, y también afecta la longevidad de las cerdas. Koketsu et al, encontraron evidencia de que agregar fibra a las dietas de gestación puede mejorar la longevidad de las cerdas.
GRASA DORSAL Y CONDICIÓN CORPORAL
El grosor de la grasa dorsal (GD) es importante en primerizas y cerdas primíparas, ya que está relacionado con la longevidad de la cerda. Algunos autores sugieren que el rango ideal de GD de las primerizas estaría entre 16 y 20mm, aunque este rango puede variar y está claramente influenciado por la genética de la cerda. Flisar et al, encontraron que las primerizas con grasa dorsal más gruesa tenían camadas más pequeñas en los 3 primeros partos. Las cerdas con grasa dorsal 10mm más gruesa parieron más camadas (0,41 en promedio) a lo largo de su vida y fueron sacrificadas 50 días después.
Farmer et al, encontraron beneficioso para las cerdas primíparas tener una mayor BF (es decir, de 20 a 26mm) al final de la gestación para lograr un desarrollo mamario óptimo y una mayor ganancia de peso corporal de la camada en la lactancia posterior. Los resultados indican que una mayor GD en la gestación tardía de las cerdas primíparas tiende a aumentar la ganancia de peso de la camada debido a una mayor producción de leche posiblemente relacionada con un mejor desarrollo y preparación de las glándulas mamarias. Dado que la mejora en la ganancia de peso de los lechones fue modesta (8,5%), las cerdas más gordas perdieron más GD para el mismo peso vivo de los lechones, y que la correlación más fuerte entre GD y los parámetros medidos en la ubre se produjo con tejido no parenquimatoso, se recomienda mantener cerdas primíparas al final de la gestación en un rango de GD entre 15 y 26mm.
El objetivo principal en la parte final de la crianza es alentar la expresión temprana del estro puberal y aparear con éxito a las primerizas mientras continúan creciendo hacia su tamaño corporal maduro. Varias estrategias son posibles. El enfoque específico puede variar de una granja a otra dependiendo de la genética y las prácticas de manejo. Aunque se ha demostrado que las restricciones severas de proteínas o la ingesta desequilibrada de aminoácidos esenciales retrasan el inicio de la pubertad, la restricción moderada de proteínas durante el período de crianza no parece influir en la edad del 1º estro en las nulíparas. La literatura más antigua indica que las primerizas de reemplazo seleccionadas deben recibir un límite de energía de alimentación de 100 a 104kg de peso corporal o hasta 2 semanas antes del apareamiento para que no engorden demasiado. No obstante, Foxcroft et al, Williams et al y Gill, presentaron evidencia de que la gordura no es un problema con las hembras modernas de genotipo magro de línea materna, que depositan y movilizan tejido magro con poco impacto en los depósitos de tejido graso. El desarrollo de genotipos ultradelgados ha tenido efectos negativos sobre la longevidad y la productividad de por vida de las primerizas de reemplazo. Esto ha llevado a la necesidad de mejorar y conservar la gordura en las nulíparas al alimentarlas con una dieta baja en proteínas (11,3% de proteína cruda, 0,45% de lisina, 13MJ de energía digestible/kg) antes y durante la gestación para restringir la ganancia magra y aumentar la deposición de grasa. En genotipos de magro medio o bajo, las nulíparas tenderán a consumir más energía de la necesaria para lograr una condición corporal ideal, por lo que se volverán demasiado gordas. Por lo tanto, se recomienda limitar la alimentación con esos genotipos después de que haya ocurrido la selección.
Gill, encontró que los aumentos en la gordura logrados por la dieta durante la crianza son transitorios. Cualquier efecto residual había desaparecido cuando se destetó la 1ª camada. Los posibles beneficios de protección para la longevidad de las cerdas al alimentarlas con una dieta baja en proteínas durante la crianza de primerizas probablemente se deban a la reducción a largo plazo del peso corporal de las cerdas y, a su vez, a la reducción del riesgo de lesiones en patas y pezuñas. Un enfoque más holístico sería considerar cómo mejorar el bienestar general y la forma física de primerizas y cerdas.
CONCLUSIÓN
La nutrición durante la crianza de primerizas juega un papel importante, ya que puede afectar la tasa de crecimiento, la condición corporal óptima, el inicio temprano del celo, el desarrollo del tracto reproductivo y las glándulas mamarias, y la buena condición de las extremidades. Es importante centrarse en el bienestar y la forma física y crear buenas condiciones ambientales desde el momento del nacimiento de una primeriza y continuar durante la crianza. Dado que las fallas reproductivas son la causa más común de sacrificio, sería apropiado investigar más a fondo el efecto de la nutrición y la estrategia de alimentación en el desarrollo y la funcionalidad del tracto reproductivo durante la crianza y su relación con el rendimiento de por vida de la cerda. Debido al mayor número de lechones nacidos por camada, también es apropiado enfocarse en una estrategia nutricional que potencie el desarrollo mamario para lograr una mayor producción de leche durante la lactancia.
Fuente: Anaporc
Deja un comentario