

La insuficiente producción de materias primas proteicas de la Unión Europea ha hecho de ella el principal importador mundial de harina de soja (HS) con 20,1 millones de toneladas anuales. La situación es más preocupante en España, con una dependencia más elevada, ya que cada año se consumen 4,47 millones toneladas teniendo que ser la mayor parte importadas (CESFAC, 2012). Una forma de minimizar el riesgo que supone el elevado coste y la disponibilidad de la soja es la utilización de fuentes proteicas alternativas.
En este sentido, el guisante presenta una composición nutricional interesante para el ganado porcino, con un porcentaje de almidón cercano al de los cereales, fibra de buena calidad y un contenido en proteína bruta (PB) que varía del 14% al 33% dependiendo de la variedad, la época de siembra y la técnica de cultivo (Castell y Guenter, 1996; González-García, 2001). Su proporción en lisina, treonina y triptófano es considerable pero es deficitario en aminoácidos azufrados, si consideramos el criterio de proteína ideal para porcino (FEDNA, 2013). Hoy en día, las llamadas variedades de primavera son las que se emplean mayoritariamente en la fabricación de piensos, al parecer porque presentan un contenido adecuado de PB y más favorable de factores antinutritivos (FEDNA, 2010).
La inclusión de elevados niveles de guisantes en piensos se ha limitado por la presencia de dichos factores antinutritivos, principalmente inhibidores de proteasas (IP), tanto de tripsina (IT) como de quimotripsina (IQ), que dificultan la actividad de las enzimas proteolíticas reduciendo la digestibilidad de la proteína (Guillamón et al., 2012; Muzquiz, 2012). Estos efectos son más acusados en animales jóvenes ya que se ha observado que la tolerancia a los factores antinutritivos aumenta con la edad (Bengala-Freire et al., 1989; Mateos et al., 2008). Al tratarse de sustancias termolábiles, los IP se pueden inactivar mediante tratamientos físicos, como la extrusión húmeda, que puede reducirlos en un 94% (Marlier et al., 1989; Adamidou et al., 2011). Su resistencia depende de la naturaleza química, siendo más estables los IP de Bowman-Birk presentes en los guisantes, y menos los de Kunitz, que se encuentran, también y en mayor proporción, en la soja (Birk, 1985; Van Amerongen et al., 1998). Sin embargo, estos tratamientos, que resultan más interesantes en lechones y con guisantes de elevada actividad inhibitoria, suponen un encarecimiento del pienso y, si son demasiado agresivos, pueden reducir la digestibilidad de los aminoácidos, principalmente de la lisina, que es sensible a las altas temperaturas (Crevieu-Gabriel, 1999).
El objetivo del ensayo fue la sustitución de la HS y el haba de soja extrusionada por 3 variedades distintas de guisantes de invierno (Cartouche, Iceberg y Luna) en dietas de cerdos de engorde (22-125kg de peso vivo, PV), lo que supuso un incremento de los niveles de IP en el pienso, y evaluar su efecto sobre los rendimientos productivos y la calidad de canal y la carne.
Material y métodos
Animales e instalaciones experimentales
Todos los procedimientos empleados en el ensayo cumplieron la normativa sobre protección de los animales utilizados para experimentación y otros fines científicos reflejada en el Boletín Oficial del Estado (2013). Se emplearon 192 cerdos de 61 días de edad y 22,10 ±2,74kg PV, de cruce híbrido Duroc x (Landrace x Large white), todos machos castrados a los 5 ±3 días de edad, procedentes de una granja situada en San Pedro Manrique (Soria, España). Al comienzo del ensayo, todos los cerdos fueron identificados y pesados individualmente, asignándose a los diferentes tratamientos experimentales en función del PV inicial. El ensayo se llevó a cabo en las instalaciones del Centro de Pruebas de Porcino del Instituto Tecnológico Agrario (ITACyL, Consejería de Agricultura y Ganadería de la Junta de Castilla y León), localizado en Hontalbilla (Segovia, España). Los cerdos fueron alojados en 4 salas, con 12 departamentos por sala y 4 animales por departamento (1,40m2 por cerdo), con cama de paja, tolva tipo holandés y un bebedero de chupete. Las condiciones ambientales se controlaron automáticamente durante todo el periodo experimental, siendo la temperatura de 20 ±2ºC y la humedad relativa de 50 ±20%.
Diseño experimental y piensos
El diseño fue en bloques completos al azar, con 4 tratamientos experimentales en función de la cantidad de IP (o variedad de guisante) incluida en el pienso: control con HS (CON), guisante Cartouche (dieta CAR), guisante Iceberg (dieta ICE) y guisante Luna (dieta LUN). Estas variedades de guisante tienen elevado interés agronómico, con un buen rendimiento por hectárea, que puede incluso superar a los cereales, y están muy adaptadas a las condiciones de Castilla-León y Castilla La Mancha (Ney y Duc, 1997; Caminero, 2002).
Hubo 4 bloques, con 3 réplicas por tratamiento en cada uno y 4 cerdos por réplica. Para los resultados productivos y características de la canal la unidad experimental fue la réplica, y para la grasa intramuscular y perfil de ácidos grasos fue la canal. De 12 canales por tratamiento, elegidas al azar (una por réplica), se obtuvieron una muestra de lomo y otra de grasa subcutánea, a nivel de la última costilla (longissimus thoracis) y final del sacro (semimembranosus), respectivamente, con el fin de estudiar el efecto del tratamiento alimenticio sobre el porcentaje de grasa intramuscular y el perfil de los ácidos grasos principales.
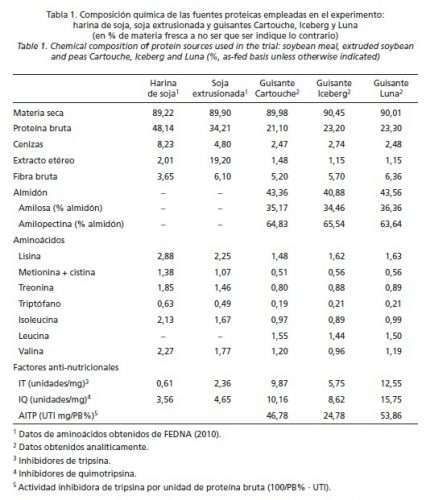
La Tabla 1 muestra la composición química analizada de las variedades de guisantes empleados en el experimento.
Los piensos fueron fabricados en Soria, y se formularon de acuerdo con FEDNA (2010) para cumplir o exceder los requerimientos nutricionales en cerdos de esa edad (FEDNA, 2013). Se administraron ad libitum en forma de gránulo de 3 mm.
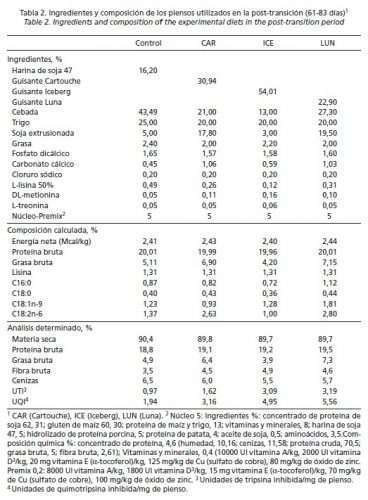
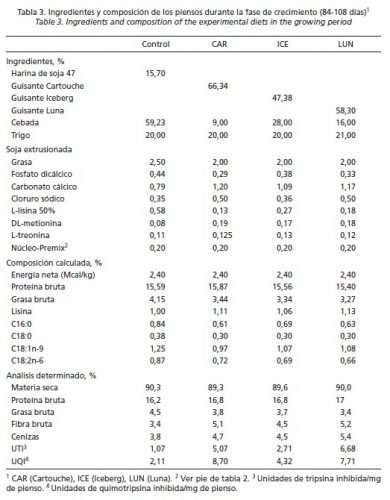
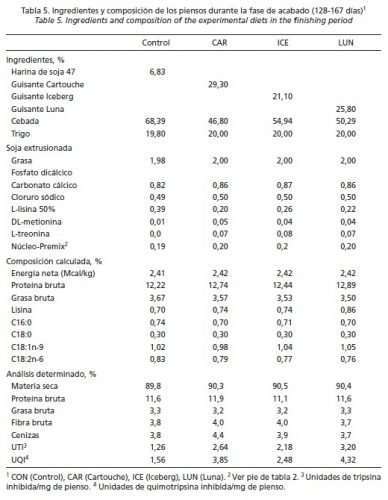
En la fase estárter (61-83 días de edad), no se sustituyó la soja por completo, utilizándose soja extrusionada como fuente de proteína y energía en los piensos con guisantes. La idea era no sobrepasar niveles de IT superiores a 4 UTI/mg de pienso, tolerables a esa edad (Batterham et al., 1993). Por este motivo, y para determinar la digestibilidad (ileal estandarizada) de proteína y aminoácidos (AA) limitantes de los distintos guisantes, se aplicaron las ecuaciones predictivas de Grosjean et al. (2000). En ellas se tiene en cuenta la actividad inhibidora de tripsina por unidad de proteína bruta (fórmula) para cada uno de los siguientes nutrientes: PB (-0,1975 · AITP + 84,72), Lisina (-0,1617 · AITP + 87,84), Metionina (-0,2630 · AITP + 86,46), Cistina (-0,2029 · AITP + 78,55), Treonina (-0,2171 · AITP + 82,61) y Triptófano (-0,3536 · AITP + 80,44) (Tabla 1). De esta manera se facilitó la formulación de los piensos con guisantes. Sin embargo, los niveles de IT y de IQ fueron más altos en los piensos que contenían guisantes Iceberg y Luna en post-transición, y a partir de crecimiento con Cartouche y Luna. Los ingredientes y los análisis de los piensos experimentales se muestran en las Tablas 2, 3, 4 y 5.

Rendimientos productivos y calidad de la canal
Diariamente se supervisó el estado sanitario de los cerdos en la granja, no registrándose incidencias ni mortalidad que pudieran afectar a los resultados del ensayo. El peso de los animales se recogió de forma individual a los 61, 83, 108, 127 y 167 días de edad mediante una báscula digital. Asimismo, se controló el aporte y el rechazo de pienso entre los controles de peso. Con todos estos datos se calcularon, por departamento, la ganancia media diaria (GMD), el consumo medio diario (CMD) y el índice de conversión (IC) para cada periodo: post-transición (61-83 días), crecimiento (84-108 días),
engorde (109-127 días) y acabado (128-167 días de edad). Tras el control de peso y de ingestión de alimento del último día, se midió el espesor de tocino dorsal mediante un equipo de ultrasonidos (Piglot 105, SFK Technology, Dinamarca) que también proporcionó el porcentaje de magro. Para ello se realizaron 2 medidas: a nivel de la 3ª-4ª últimas costillas y entre la 3ª-4ª vértebras lumbares, en ambos casos a 8cm de la línea media y repitiendo en los 2 flancos del animal.
El sacrificio, con una media de 121kg PV, se llevó a cabo en un matadero industrial de Cuenca. Previo al sacrificio, los animales se mantuvieron en ayuno 12 horas pero con agua a libre disposición. El aturdimiento se efectuó en cámara de dióxido de carbono, con una concentración mínima del 80% durante 45 segundos. Tras el desangrado, escaldado y eviscerado se tomó el peso de la canal en caliente para calcular su rendimiento. A las 24 horas postmortem se llevó a cabo el despiece de todas las canales y, de cada media canal izquierda, se registró el peso individual de las principales piezas nobles (jamones, paletas y chuleteros) tras el perfilado, para calcular su rendimiento.
Análisis de laboratorio
La composición química de los piensos y de los guisantes fue determinada en el laboratorio de I+D Agroalimentario del ITACYL (finca Zamadueñas, Valladolid), y en el Departamento de Tecnología de los Alimentos, Sección de Tecnología Vegetal, del Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (INIA, Madrid), de acuerdo con los procedimientos descritos en el Diario Oficial de la Unión Europea (2009). La concentración de AA en guisantes se determinó mediante cromatografía de intercambio iónico (Hewlett-Packard 1100, Waldbronn, Alemania) después de hidrólisis ácida, aplicando el procedimiento descrito por Jones et al. (1981). Para valorar las concentraciones de metionina y cistina, muestras separadas se oxidaron con ácido perfórmico antes de la hidrólisis midiéndose como sulfona metionina y ácido cisteico, respectivamente (Moore, 1963). El triptófano se calculó mediante cromatografía líquida de alta resolución con fluorescencia, después de hidrólisis alcalina (hidróxido de bario) durante 20 horas a 110ºC (DOUE, 2009).
La actividad IT (UTI/mg de muestra; guisantes, soja y piensos), fue valorada con el método de Kakade et al. (1974) modificado por Grant et al. (1995) y Muzquiz et al. (2004). Para ello, se tomaron muestras por duplicado (0,025gr) que se molieron usando un molino (Retsch modelo Z-I, Sttutgart, Alemania) provisto de una criba de 0,5mm, para posteriormente, ser extraída con 1ml de HCl 0,05 molar, mediante un agitador orbital durante 1 hora, a una temperatura de 5ºC. El extracto se centrifugó a 11.000 x g durante 4min, y el sobrenadante se midió para el cálculo de la actividad, usando N-α-Benzoil-DL-Arginina-pnitroanilida como sustrato específico. La actividad IT se calculó por lectura de absorbancia contra un blanco a 410nm, usando un espectrofotómetro ultravioleta (modelo DU-7, Beckmann Instruments Inc., Fullerton, EE.UU.).
En el caso de la actividad IQ (UQI/mg muestra) se empleó el procedimiento de Sathe y Salunkhe (1981). Las muestras se procesaron del mismo modo que para la actividad IT y con una solución tampón (100mM Tris-HCl conteniendo 20mM de CaCl2 y pH 7,8) a 30ºC con N-Benzoil-L-Tirosina-Etil Ester como sustrato. La absorbancia se midió 7 minutos después de la adición del sustrato a 256nm con el equipo indicado para la actividad IT.
La grasa intramuscular del lomo fue extraída mediante el método propuesto por Marmer y Maxwell (1981). La grasa subcutánea se extrajo mediante el procedimiento de Bligh y Dyer (1959) y se metiló con la técnica de Sandler y Karo (1992). La composición de los ácidos grasos de la grasa subcutánea obtenida se determinó por cromatografía de gases en las siguientes condiciones: temperatura del horno a 170ºC, temperatura del inyector a 250ºC, temperatura del detector a 250ºC, flujo de gas portador a 2,5mL/min ºC split 1/50. La identificación de los ácidos grasos se realizó por inyección de ésteres metílicos de patrones y su comparación posterior con los tiempos de retención de los ésteres metílicos de los ácidos grasos (BOE, 2004). El cromatógrafo utilizado fue un Hewlett Packard – HP-5890 serie II; el sistema de inyección empleado fue split con un detector de ionización de llama. La columna fue HP-innowax capilar con una longitud de 30m, diámetro interno de 0,32mm y un grosor de fase de 0,25µm. Los ácidos grasos se expresaron como porcentaje (gramos de ácido graso por 100gr de ácidos grasos).
Análisis estadístico
El diseño fue en bloques completos al azar, utilizándose los procedimientos GLM y REG del programa estadístico SAS (2004), siendo el departamento con 4 cerdos la unidad experimental (réplica; 12 por tratamiento). Para la grasa intramuscular y perfil de ácidos grasos la unidad experimental fue la canal (réplica; 12 por tratamiento). El modelo fue el siguiente: (fórmula) donde, Y: CMD, GMD, IC, PESO, UTI, UQI, porcentaje magro de canal, espesor de tocino dorsal, así como pesos y rendimientos de canal, jamón, paleta y chuletero en los datos obtenidos en el matadero, porcentaje de grasa intramuscular del lomo y proporción de los principales ácidos grasos de la grasa subcutánea; µ: media general; pienso: CON, CAR, ICE y LUN; α: coeficiente de regresión parcial entre peso inicial (P1) e Y; P1: covariable, peso al inicio del periodo experimental y ε: error residual.
El efecto del bloque (sala) se retiró del modelo al no ser significativo (P > 0,05). Las medias se compararon y separaron mediante un t-test, siendo P < 0,05 clasificado como la diferencia significativa y P < 0,10 tendencia. Los datos se presentan en tablas como medias corregidas por mínimos cuadrados. Se obtuvieron, igualmente, ecuaciones de regresión múltiple que relacionaron CMD, GMD e IC con la ingesta diaria de IT, IQ y la ratio IT/IQ en las distintas fases del cebo. La covariable P1 fue retirada del modelo estadístico para el estudio de las características de la canal. Del mismo modo, se llevó a cabo un análisis estadístico adicional que consideró el peso final de los cerdos en el estudio de la canal. Resultados Análisis de ingredientes y piensos La Tabla 1 muestra la composición química de las fuentes proteicas usadas en el ensayo. La proteína bruta superó el 20% en las 3 variedades de guisante. La variedad Cartouche mostró el mayor porcentaje de extracto etéreo y el menor de lisina mientras que la variedad Iceberg tuvo las menores proporciones de almidón y de valina. Los resultados analíticos diferenciaron los guisantes por su actividad antitrípsica del siguiente modo: Luna > Cartouche > Iceberg; los IQ superaron a los IT en las 3 variedades pero en diferentes proporciones (2,9, 49,9 y 25,5%, para Cartouche, Iceberg y Luna, respectivamente).
Las Tablas 2, 3, 4 y 5 reflejan la composición de las dietas usadas en el ensayo. La actividad de IT en los piensos post-transición (61-83 días de edad) fue inferior a 4 unidades/mg en todos los casos, siendo la dieta LUN la que presentó mayor capacidad inhibidora cumpliendo así con el planteamiento inicial. Durante la fase de crecimiento (94-108 días de edad), el nivel de inclusión de guisantes en los piensos fue muy elevado y los IP alcanzaron, en general, mayores valores que en la fase anterior debido a que se sustituyó por completo la soja extrusionada y, por otro lado, el porcentaje de proteína en la dieta seguía siendo alto. En las fases de engorde (109-127 días de edad) y acabado (128-167 días de edad), la actividad inhibidora disminuyó de manera notable en las 3 dietas con guisantes; sin embargo, en el pienso CON la actividad varió menos y permaneció más constante durante todo el periodo experimental (61 a 167 días de edad).
Resultados productivos, calidad de canal y carne
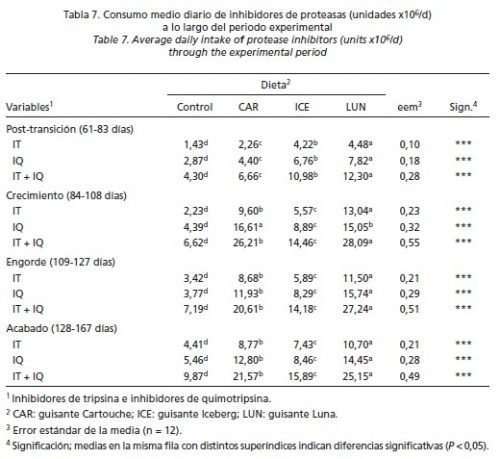
En la Tabla 6 se muestra el efecto del contenido en IP (y de la variedad de guisante) en los rendimientos productivos de los animales. Durante el periodo de post-transición (61-83 días de edad), no se detectaron diferencias para CMD ni GMD pero los cerdos que consumieron la dieta CAR presentaron mejor IC que los que comieron la dieta ICE, con los otros tratamientos en una posición intermedia (P < 0,01). Durante el periodo de crecimiento (84-108 días de edad), las dietas CON e ICE dieron mayores crecimientos (P < 0,001) e ingestiones de pienso (P < 0,05) y mejores conversiones alimenticias (P < 0,001) que las dietas CAR y LUN. Estos efectos comenzaron a diluirse durante la fase de engorde (109-127 días de edad), no detectándose diferencias en GMD ni IC pero sí en CMD, presentando los cerdos con las dietas CAR y LUN menores consumos que los de la dieta CON (P < 0,05). Durante la fase de acabado (128-167 días de edad), no se encontró influencia alguna de la dieta sobre ninguna variable estudiada. En la fase global (61-167 días de edad), los animales que recibieron la dieta CON comieron más pienso (P < 0,05), crecieron más rápido (P < 0,01) y fueron más pesados al sacrificio (P < 0,01) que los cerdos de las dietas CAR y LUN, con aquellos que ingirieron la dieta ICE en una posición intermedia. La ingestión media diaria de IQ fue mayor que la de IT a lo largo de todo el periodo experimental (Tabla 7). Independientemente de la fase estudiada, el consumo de IP (tanto de IT como de IQ) fue mayor con las dietas que incluían guisantes que con la dieta CON; y entre las dietas con guisantes resultó mayor con la dieta LUN que con la dieta CAR ocupando la dieta ICE una posición intermedia (P > 0,05). Por fases, se observó que los cerdos que comieron la dieta CON aumentaron progresivamente la ingestión de IP a lo largo del ensayo. Sin embargo, con todas las dietas que incluían guisantes, el consumo de IP aumentó notablemente de la fase de post-transición a la fase de crecimiento; se redujo posteriormente en la fase de engorde y se mantuvo durante la de acabado (resultados medios: 10 vs 22,9 vs 20,7 vs 20,9 x 106 unidades de IP/día, respectivamente).
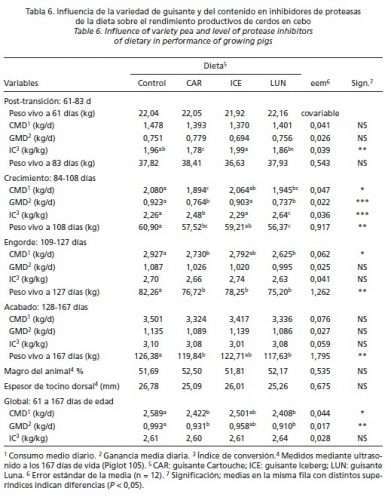
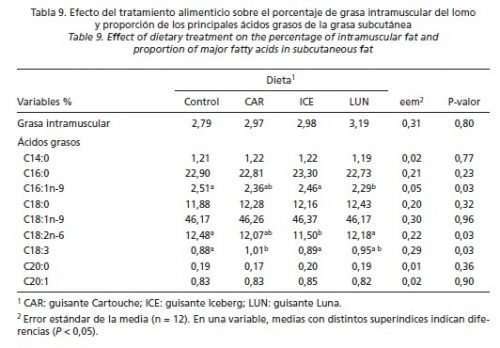
En las ecuaciones de regresión obtenidas relacionando las variables productivas con la ingesta diaria de IP tan sólo se obtuvieron resultados significativos en el CMD (R2: 0,43) y la GMD (R2: 0,51) en la fase de post-transición (P < 0,001), y en la GMD (R2: 0,39) y el IC (R2: 0,49) en la fase de crecimiento (P < 0,001). No se detectó influencia de la dieta sobre ninguno de los parámetros de calidad de canal evaluados, ni en la grasa infiltrada del lomo ni en el porcentaje de los principales ácidos grasos (C16:0, C18:0, C18:1n-9 (P > 0,05); sin embargo, sí que hubo diferencias en los porcentajes de C16:1n-9, C18:2n-6 y C18:3 (P < 0,05) (Tablas 8 y 9).
Discusión
El guisante es la proteaginosa más estudiada y mejorada en los últimos 30 años. Las variedades tolerantes al frío, sembradas en octubre-noviembre y llamadas guisantes de invierno no han tenido el protagonismo que merecen, siendo las variedades de primavera las más utilizadas en los piensos al relacionarse con mayores rendimientos por hectárea, más porcentaje de PB y menores cantidades de IP, a priori el factor limitante para su inclusión en piensos. Esto ha sucedido por el impulso de los países más productores y consumidores de guisante proteaginoso, como es el caso de Francia y, en nuestro caso, por el desconocimiento, al sembrar variedades que no se adaptan a las condiciones edafoclimáticas (Caminero, 2002). En Castilla y León, principal zona productora de España, las variedades mejoradas de invierno ofrecen resultados muy diferentes y superan a las llamadas de primavera: más producción por hectárea, porcentaje de PB superior en 1 o 2 puntos y no siempre más cantidad de IP, al no ser éste un carácter ligado a la época de siembra o resistencia al frío (García-González, 2001; Caminero, 2002). De hecho, las 3 variedades de guisantes de invierno empleadas en el presente experimento presentaron contenidos en proteína bruta superior al 20,6% publicado por FEDNA (2010) para las variedades de primavera. De acuerdo con Carrouée et al. (1994), y teniendo en cuenta los resultados de IT, los guisantes testados se podrían clasificar del siguiente modo: actividad baja para Iceberg (4 a 7 unidades de IT/mg), media para Cartouche (7 a 10 unidades IT/mg) y alta para Luna (10 a 13 unidades IT/mg). Los IQ superaron a los IT en las 3 variedades, pero en diferente medida, llegando al 50% en el caso de Iceberg, sin que exista una relación lineal entre ambos inhibidores. Posiblemente éste sea un carácter ligado a la variedad del guisante.
Rendimiento productivo. Calidad de canal y piezas nobles
Hay numerosos trabajos valorando dietas que sustituyen parcial o totalmente soja por guisante en cerdos durante el cebo, aunque utilizan fundamentalmente guisantes de primavera, y la mayoría no tienen en consideración los niveles de IT y menos aún los IQ, tanto en el guisante como en la dieta. Los resultados son muy dispares; así, Castell y Guenter (1996) limitaron la incorporación de guisante en las dieta de engorde y acabado a 20 y 35%, Mathé et al. (2003) llegan al 39% (guisante con 2,47 UTI/mg), mientras Stein et al. (2004) indicaron un máximo del 36%. Con la información disponible es de suponer que se trata de guisantes con baja actividad IP y en todos los casos de primavera. Otros trabajos reemplazan completamente la soja por guisantes. Así, Gatel et al. (1989) obtuvieron el mismo rendimiento en cerdos alimentados con un pienso único durante la fase de cebo (28 a 101kg) que tenía un 41% de guisante de primavera variedad Finale (2,76 UTI/mg) sin soja frente a un control de soja y trigo. Mientras, Stein et al. (2006) en cerdos de 22 a 123kg no observaron diferencias en rendimiento productivo ni en calidad de la canal al suministrar durante las distintas fases de cebo 66, 48 y 36% de guisante primavera sin soja frente a una dieta con soja y cereal. Son situaciones similares al presente trabajo, incluso eliminando la soja a edades más tempranas, pero con guisantes primavera y, se supone, bajos en IT.
Doti et al. (2014) compararon 2 dietas isoenergéticas en cerdos de 63 a 93kg (una con 78% de cebada y 19% de HS y otra con 49% de cebada, 40% de guisantes de primavera y 6% de HS), que contenían 17,3 y 18% de proteína bruta, respectivamente, no detectando diferencias en parámetros productivos ni tampoco en la digestibilidad de la proteína.
La utilización de distintas variedades en piensos de cebo quedan reflejadas en los estudios de Grosjean et al.(1989), que en un primer ensayo compararon 3 dietas: control con 21% de HS, 30% guisante de invierno Frisson (8,9 UTI/mg) + 7% HS y 30% de guisante primavera Finale (4,1 UTI/mg) + 6HS. No hubo diferencias entre los cerdos (30 a 100kg) que comieron los guisantes y el control, pero sí entre las dietas con los guisantes, con menor CMD y GMD de los animales alimentados con la dieta del guisante Frisson (2,290 vs 2,340kg y 0,757 vs 0,794kg, de CMD y GMD para Frisson y Finale, respectivamente; P < 0,05). La actividad IT del pienso con Frisson fue de 2,7 y 1,3 UTI/mg para la dieta del guisante Finale. En un segundo ensayo con el mismo tipo de cerdos y 2,4 y 1,6 UTI/mg en el pienso con Frisson (27%) y Finale (27%) respectivamente, y 5,5% de HS en ambos, no hubo diferencias entre los cerdos alimentados con los dos tipos de guisantes (P > 0,05). Son cantidades de IT en el pienso con la variedad Frisson similares a las del pienso ICE, sin embargo no sustituyen por completo la soja, son dietas únicas y, prácticamente, con un mismo diseño, se obtienen diferentes resultados.
La edad de inicio del consumo de guisante parece influir en el rendimiento posterior, así Bengala-Freire, et al. (1989) en cerdos desde 27 a 102kg vieron que un porcentaje de guisante superior al 30%, con 2,85 UTI/mg, disminuyó el CMD y la GMD. Por el contrario, Newman et al. (2011) reemplazaron la soja por derivados de guisante seco (“chips pea”): 45% a partir de los 57kg y 30% desde 93 a 127kg, sin diferencias significativas en relación con el control de soja y maíz. En cerdos pesados (40 a 158kg), Prandini et al. (2011) sustituyeron la soja por guisante sin observar efectos negativos en rendimiento ni en calidad de la canal.
Los resultados de las Tablas 6 y 7 muestran que los cerdos se adaptan a consumos elevados de IP según aumenta la edad. Concretamente, en la fase de acabado no se presentaron diferencias productivas entre los animales de los 4 tratamientos (P > 0,05). En una experiencia parecida Njoka (2008) enfrentó 4 dietas durante la fase de acabado (80 a 123kg), una control de soja con 3 sin soja y que incorporaban un 30% de guisantes, cada una de un tipo: de invierno, de verano y de primavera. Sólo observó una tendencia en el consumo, siendo mayor en los animales alimentados con las dietas invierno y verano que con las primavera y control: 4 y 3,8 vs 3,5 y 3,4kg, respectivamente. En un ensayo con la variedad Cartouche (9,87 UTI y 10,16 UQI), pero sin periodo de adaptación (post-estárter) y sustituyendo a la soja a partir de los 30kg de peso hasta los 125kg, de Mercado et al. (2013) y Gómez-Fernández et al. (2013) no obtuvieron diferencias al compararlo con un pienso control de soja, ni en producción ni en calidad de la canal.
En las experiencias anteriores, los guisantes, con la excepción de la variedades Frisson y Cartouche (invierno), y los piensos, tuvieron unos niveles de IT muy inferiores a los del presente ensayo. A pesar de ello hubo diferencias debidas a consumos menores. Incluso en la fase estárter (Tabla 6), y duplicando la cantidad de IP del CON, los animales que consumieron ICE, tuvieron el mismo resultado para CMD, GMD e IC. Es un resultado similar al de Batterham et al. (1993) que compararon una dieta sin guisante, con garbanzos (Cicer arietinum) y guandú (Cajanus cajan), frente a un control de soja, con cerdos desde 20 a 50kg, sin encontrar diferencias productivas con cantidades de 4,7 UTI/mg y 4,5 UQI/mg en los piensos.
La posible reducción del consumo, que refieren diferentes autores, puede deberse a la presencia de IP o a otras causas. McLaughlin et al. (1983) lo relacionan con niveles elevados de IT que pueden activar la liberación de colecistoquinina (CCK) hormona relacionada con la saciedad; para Grosjean et al. (1997) los IT disminuirían la digestibilidad del triptófano, AA con efecto sobre el apetito. Jaikaran et al. (2002) en un ensayo con guisantes de distinto color (amarillos y verdes) y cerdos de más de 25kg, no encontraron diferencias en CMD al compararlos con una dieta control de soja (P > 0,05). Sin embargo, la presencia de saponinas en los guisantes, que según la variedad y el tipo de saponina (B o DDMP) varía de 0 a 1,5g/kg, podría reducir el consumo debido al sabor amargo (Heng et al., 2006), aunque Price et al. (1985) indican que el rechazo parcial de la dieta empezaría a partir de 2g/kg de saponina.
El efecto del porcentaje y tipo de almidón es una cuestión a tener en cuenta. La cantidad (%) fue muy similar en las distintas dietas según fases: 37 ± 1% en Post-transición, 39 ± 2% en Crecimiento, 46 ± 1% en Engorde y 49 ± 1% en Acabado. Es cierto que en legumbres como los guisantes la proporción de amilosa es mayor que en los cereales, lo que puede variar la degradación del almidón, el índice glucémico, y hasta cierto punto la ingesta. Así lo refieren autores como Bengala-Freire, et al. (1989), aunque Doti et al. (2014) no encontraron diferencias con distintas fuentes de almidón (muy o poco resistentes a la degradación). En el presente estudio, si bien varía la fuente y tipo de almidón según inclusión de guisantes y cebada, la disminución del consumo se relaciona más con la ingesta elevada de inhibidores que con el tipo de almidón (Tabla 5). Sirva como ejemplo el CMD en la fase de Post-transición, no significativo a pesar de los altos niveles de almidón de guisante en el pienso ICE en relación con el CON.
Las diferencias de rendimiento en los resultados, se relacionaron directamente con un menor consumo de pienso, ligado a la edad de los cerdos y a la cantidad de IP que comieron, y no se manifestaron en el espesor de tocino dorsal, porcentaje magro, ni en los datos de la canal o piezas nobles.
Grasa intramuscular del lomo y ácidos grasos de la grasa subcutánea
Como cabía esperar, debido a que el pienso control y los experimentales administrados durante las fases de engorde y acabado fueron isoenergéticos, isoproteicos y tuvieron análogo tenor graso, el tratamiento alimenticio no tuvo influencia sobre el porcentaje de grasa intramuscular (López-Bote y Rey, 2004). Así mismo, debido a que la composición calculada de los piensos de engorde y acabado en los principales ácidos grasos, y el consumo medio diario de pienso fueron similares según tratamiento, tampoco eran de esperar variaciones en el perfil de ácidos grasos de la grasa subcutánea. Sin embargo, y dentro de los ácidos grasos mayoritarios, la ligera mayor proporción de ácido linoleico (C18:2n-6) obtenida en los cerdos que consumieron el pienso CON (solo significativamente mayor que los cerdos que comieron el pienso ICE), puede explicarse por un contenido más elevado de ácido linoleico de este pienso durante las fases de engorde y acabado, y por un mayor consumo, aritméticamente superior, del pienso indicado.
Las diferencias observadas de ácido linoleico, entre los tratamientos en los que se sustituyó soja y cebada por guisantes, no tienen una explicación coherente, salvo que sean debidas a efectos individuales inherentes a los animales que fueron elegidos al azar para el posterior análisis de ácidos grasos.
Podemos concluir que la edad de los cerdos influye en la tolerancia a los niveles de IP derivados de un pienso con guisante. En fases iniciales, son capaces de soportar cantidades de IP que pueden triplicar las de un pienso control de soja y, en las fases de engorde y acabado, incluso más elevadas, sin mermas productivas ni de calidad de canal o piezas nobles, ni del contenido de grasa intramuscular en el lomo y proporción de ácidos grasos principales (C16:0, C18:0, C18:1n-9) en la grasa subcutánea. Los resultados sugieren que los guisantes de variedades de invierno suponen una alternativa a la soja perfectamente viable si se adaptan, según su contenido en IP, a las diferentes fases productivas.
Autores
- E. Gómez-Izquierdo1, E. de Mercado, J. Gómez-Fernández, C. Tomás. Centro de Pruebas de Porcino del Instituto Tecnológico Agrario de Castilla y León. Consejería de Agricultura y Ganadería. Hontalbilla, Segovia, España.
- E. Guillamón. Centro para la Calidad de los Alimentos, INIA, Campus Universitario Duques de Soria, Soria, España.
- A. Varela, M. Muzquiz, M.M. Pedrosa. Departamento de Tecnología de los Alimentos, INIA, Madrid, España
- P. López-Nuez. Copiso, Soria, España.
- M.A. Latorre. Departamento de Producción Animal y Ciencia de los Alimentos, Facultad de Veterinaria, Instituto Agroalimentario de Aragón-Universidad de Zaragoza (IA2-UZ), Zaragoza, España.
Bibliografía
- Adamidou S, Nengas I, Grigorakis K, Nikolopoulou D, Jauncey K (2011). Chemical composition and antinutritional factors of field peas (Pisum sativum), chickpeas (Cicer arietinum), and faba beans (Vicia faba) as affected by extrusion preconditioning and drying temperatures. Cereal Chemistry Journal 88: 80-86.
- Batterham ES, Saini HS, Andersen LM, Baigent RD (1993). Tolerance of growing pigs to trypsin and chymotrypsin inhibitors in chickpeas (Cicer arietinum) and pigeonpeas (Cajanus cajan). Journal of Science Food and Agriculture 61: 211-216.
- Bengala-Freire J, Hulin JC, Peiniau J, Aumaitre A (1989). Effet de la cuisson-extrusion du pois de printemps sur la digestibilité des aliments de sevrage precoce du porcelet et consequences sur les performances jusqu a l abattage. 21e Journées de la Recherche Porcine, 31-2 febrero 1989, París, Francia, pp. 75-82.
- Bligh EG, Dyer WJ (1959). A rapid method of total lipid extraction and purification. Canadian Journal of Biochemistry and Physiology 37(8): 911-917, 10.1139/o59-099.
- Birk Y (1985). The Bowman-Birk inhibitor. Trypsin and chymotrypsin inhibitor from soybean. International Journal of Peptide and Proteine Research 25: 113-131.
- Boletín Oficial del Estado (2004). Orden PRE/3844/ 2004, de 18 de noviembre, por la que se establecen los métodos oficiales de toma de muestras en canales de cerdos ibéricos y el método de análisis para la determinación de la composición de ácidos grasos de los lípidos totales del tejido adiposo subcutáneo de cerdos ibéricos.
- Boletín Oficial del Estado (2013). Real Decreto 53/2013, de 1 de febrero, por el que se establecen las normas básicas aplicables para la protección de los animales utilizados en experimentación y otros fines científicos, incluyendo la docencia. BOE 34: 11370-11421.
- Caminero C (2002). Adaptación a la siembra invernal y tolerancia al frío en guisante (Pisum sativum L.). Tesis Doctoral. Universidad de Valladolid. Instituto Tecnológico Agrario de Castilla y León (Ed).
- Carrouée B, Grosjean F, Peyronnet C, Weiss P (1994). Guisantes. Utilización en Alimentación Animal. UNIP-ITCF. París, 96 pp.
- Castell AG, Guenter W (1996). Nutritive value of peas for nonruminant diets. Animal Feed Science and Technology 60(3-4): 209-227.
- CESFAC (2012). Confederación Española de Fabricantes de Alimentos Compuestos para Animales. Mercados estadística 2011.
- Crevieu-Gabriel I (1999). Digestion des protéines végétales chez les monogastriques. Exemple des protéines de pois INRA. Production Animal 12(2): 147-161.
- de Mercado E, Gómez-Fernández J, Muzquiz M, Guillamón E, Varela A, Tomás C, López-Nuez P, Gómez-Izquierdo E (2013). Evolución del crecimiento, calidad de la canal y de piezas nobles en cerdos grasos alimentados con guisante (Pisum sativum) y alberjón (Vicia narbonensis) como fuente proteica. XV Jornadas ITEA-AIDA, 14 y 15 de mayo 2013, Zaragoza, España, vol. 1: 171-173.
- Diario Oficial de la Unión Europea (2009). Reglamento (CE) nº 152/2009 de la Comisión de 27 de enero de 2009, por el que se establecen los métodos de muestreo y análisis para el control oficial de los piensos. L 54/1-L 54/130.
- Doti S, Suárez-Belloch J, Latorre MA, Guada JA, Fondevila M (2014). Effect of dietary starch source on growth performances, digestibility and quality traits of growing pigs. Livestock Science 164: 119-127.
- FEDNA (2010). Tablas FEDNA de composición y valor nutritivo de alimentos para la fabricación de piensos compuestos. 3ª Edición (Ed. de Blas C, Mateos GG, García Rebollar P). Ed. Fundación Española para el Desarrollo de la Nutrición Animal. 502 pp. Madrid.
- FEDNA (2013). Necesidades nutricionales para ganado porcino: Normas FEDNA (Ed. de Blas C, Mateos GG, García Rebollar P). 2ª edición. Ed. Fundación Española para el Desarrollo de la Nutrición Animal.
- Gatel F, Grosjean F, Castaing J (1989). Utilisation par le porc charcutier de régimes à teneur élevée en pois de printemps (plus de 40%). 21e Journées de la Recherche Porcine, 31-2 febrero 1989, París, Francia, pp. 69-74.
- Gómez-Fernández J, de Mercado E, Muzquiz M, Guillamón E, Varela A, Tomás C, López-Nuez P, Gómez-Izquierdo E (2013). Empleo de guisante (Pisum sativum) y alberjón (Vicia narbonensis) como fuentes proteicas en dietas para cerdos grasos. Rendimiento productivo. XV Jornadas ITEA-AIDA, 14 y 15 de mayo 2013, Zaragoza, España, vol. 1: 135-137.
- González-García MR (2001). Interacción genotipo x ambiente en guisante proteaginoso (Pisum sativum L.). Tesis Doctoral. Universidad de Valladolid.
- Grant G, Dorward PM, Buchan WC, Armour JC, Pusztai A (1995). Consumption of diets containing raw soya beans (Glycine max L.), kidney beans (Phaseolus vulgaris L.), cowpeas (Vigna unguiculata L.) or lupin seeds (Lupinus angustifolius L.) by rats for up to 700 days: Effects on body composition and organ weights. British Journal of Nutrition 73(1): 17-29.
- Grosjean F, Bourdon D, Theillaud-Ricca V, Castaing J, Beague E (1989). Comparaison des pois d’hiver et de printemps dans des aliments pour porcs charcutiers présentés en farine ou en granulés. 21e Journées de la Recherche Porcine, 31-2 febrero 1989, París, Francia, pp. 75-82.
- Grosjean F, Jondreville C, Bogaert C, Bourdillon A, Peyronnet C, le Guen MP, Williatte I (1997). Utilisation d’aliments pour porcelets sevrés contenant 40% de pois. 29e Journées de la Recherche Porcine, 4-6 febrero 1997, París, Francia, pp. 197-204.
- Grosjean F, Jondreville C, Williatte-Hazouard I, Skiba F, Carrouée B, Gatel F (2000). Ileal digestibility of protein and amino acids of feed peas with different trypsin inhibitor activity in pigs. Canadian Journal of Animal Science 80: 643-652.
- Guillamón E, Mateo-Vivaracho L, Villares A, D’Arrigo M, Pedrosa MM, García-Lafuente A, Muzquiz M (2012). Inhibidores de tripsina y de quimotripsina en semillas de leguminosas. lV Jornadas de la Asociación Española de leguminosas, 6-7 Junio, Pontevedra, España, pp. 111-113.
- Heng L, Vincken JP, van Koningsveld GA, Legger A, Gruppen H, van Boekel MAJS, Roozen J, Voragen AGJ (2006). Bitterness of saponins and their content in dry peas. Journal of the Science of Food and Agriculture 86(8): 1225-1231.
- Jaikaran S, Colangelo M, William C (2002). Growth and feed intake of growing pigs fed diets containing three varieties of field peas. Advances in pork production 13: 25.
- Jones BN, Paavo S, Stein S (1981). Amino acids analysis and enzymatic sequence determination of peptides by an improved o-phthaldialdehyde precolumn labeling procedure. Journal of Liquid Chromatography 4: 565-586.
- Kakade ML, Racki JJ, McGhee JE, Puski G (1974). Determination of trypsin inhibitor activity of soy products: a collaborative analysis of an improved procedure. Cereal Chemistry Journal 51: 376-382.
- López-Bote CJ, Rey AI (2004). Efecto de la alimentación en las características del magro. Porci 84: 60-77.
- Marlier L, Focant M, Allart B, Vanbelle M (1989). Effects du floconage et de l’extrusion sur le valeur alimentaire du pois protéagineux, pour le porc charcutiere. Annales de Zootechnie 38: 237-245.
- Marmer WN, Maxwell RJ (1981). Dry column method for the quantitative extraction and simultaneous class separation of lipids from muscle tissue. Lipids 16(5): 365-71.
- Mateos GG, Valencia DG, Serrano MP, Lázaro R (2008). Las leguminosas de grano en alimentación animal: estudio del guisante. III Jornadas de la Asociación Española de Leguminosas, 2-4 de septiembre de 2008, Valladolid, España, pp. 72-94.
- Mathé D, Monéger R, Guillou D (2003). Effet du pois protéagineux sur les performances et le comportement du porc lors des transitions alimentaires. 35e Journées de la Recherche Porcine, 4-6 febrero 2003, París, Francia, pp. 127-132.
- McLaughlin CL, Peikin SR, Baile CA (1983). Trypsin inhibitor effects on food intake and weight gain in Zucker rats. Physiology and Behavior 4: 487-91.
- Moore S (1963). On determination of cystine as cisteic acid. Journal of Biological Chemistry 238: 235-237.
- Muzquiz M (2012). Componentes nutricionalmente activos en leguminosas: implicaciones en nutrición y salud. lV Jornadas de la Asociación Española de leguminosas, 6-7 Junio, Pontevedra, España, pp. 111-113.
- Muzquiz M, Welham T, Altares P, Goyoaga C, Cuadrado C, Romero R, Guillamon E, Domoney C (2004). The presence of trypsin inhibitors in Vicia faba and Cicer arietinum during germination. Journal of the Science of Food and Agriculture 84:556-560.
- Newman DJ, Harris EK, Lepper AN, Berg EP, Stein HH (2011). Effects of pea chips on pig performance, carcass quality and composition, and palatability of pork. Journal of Animal Science 89: 3132-3139.
- Ney B, Duc G (1997). The main constraints to overcome in the plant development for the winter type varieties. Grain legumes 16: 14-15.
- Njoka JG (2008). Effects of feeding Iowa-grown field peas on finishing pig performance. Graduate Theses and Dissertations. Paper 11367.
- Prandini A, Sigolo S, Morlacchini M, Cerioli C, Masoero F (2011). Pea (Pisum sativum) and faba bean (Vicia sativa L.) seeds as protein sources in growing-finishing heavy pig diets: effect on growth performance, carcass characteristics and on fresh and seasoned Parma ham quality. Italian Journal of Animal Science 10: e45.
- Price KR, Griffiths NM, Curl CL, Fenwick GR (1985). Undesirable sensory properties of the dried pea (Pisum sativum). The role of saponins. Food Chemistry 17: 105-115.
- SAS (2004). User’s Guide. SAS Institute. Cary, NC, EE.UU.
- Sandler SR, Karo W (1992). Sourcebook of advanced organic laboratory preparations. Published by San Diego u. a., Academic Press. ISBN 10: 0126185069 ISBN/13: 9780126185065.
- Sathe SK, Salunkhe DK (1981). Studies on trypsin and chymotrypsin inhibitory activities, hemagglutinating activity, and sugars in the Great Northern beans (Phaseolus vulgaris L). Journal of Food Science 46: 626-629.
- Stein HH, Benzoni G, Bohlke RA, Peters DN (2004). Assessment of the feeding value of South Dakota-grown field peas (Pisum sativum L.) for growing pigs. Journal of Animal Science 82(9): 2568-2578.
- Stein HH, Everts AKR, Sweeter KK, Peters DN, Maddock RJ, Wulf DM, Pedersen C (2006). The influence of dietary field peas (Pisum sativum L.) on pig performance, carcass quality, and the palatability of pork. Journal of Animal Science 84: 31.
- Van Amerongen A, Ostafe V, Meijer MMT, Gruppen H, Meerdink G, Bedrendsen LBJM, Koets M, Wichers JH (1998). Specific immuno-(chymo) trypsin inhibitor assays for determination of (residual) activity of Browman-Birk or Kunitz soybean trypsin inhibitors. En: Recent advances of research in antinutritional factors in legume seeds and rapeseed (Eds. Jansmanñ AJM, Hill GD, Huisman J, van der Poel AFB), EAAP publication no 93 Wageningen, Holanda. pp. 33-37
Deja un comentario