

RESUMEN
Se han controlado 250 camadas de cerdas Retinto del Andévalo que parieron en primavera y otoño. La prolificidad fue elevada y fue superior en cerdas multíparas que en primíparas. La mortalidad de lechones durante la lactancia fue superior en otoño que, en primavera, por lo que el número de lechones destetados fue más bajo en otoño. Las elevadas mortalidades al nacimiento y al destete derivadas del modelo de explotación extensivo en camping se tradujeron en unos resultados discretos al destete. De otra parte, se controlaron 164 lechones de 26 camadas incluidas en los 4 primeros partos. El peso al nacimiento de los lechones y el peso al destete fue superior en cerdas multíparas que en primíparas y, ambas variables, fueron más elevadas en los machos que en las hembras. La estación no afectó al peso al nacimiento y al crecimiento de los lechones durante la lactación.
INTRODUCCIÓN
En la comarca del Andévalo de la provincia de Huelva la explotación del cerdo Ibérico tiene una considerable importancia sobre todo bajo fórmulas extensivas o semi extensivas de producción En la formación de la línea Retinto del Andévalo han intervenido las estirpes de cerdo Ibérico Villalón y Torbiscal, siendo esta última el resultado del cruzamiento de 4 estirpes: 2 españolas (Campanario y Puebla) y 2 portuguesas (Ervideira y Caldeira). Algunos trabajos previos han sido realizados sobre caracterización reproductiva del Retinto del Andévalo (Forero et al., 2014), aunque estimamos que abundar sobre tal caracterización supone una contribución adicional nada desdeñable en el escenario científico y productivo del cerdo Ibérico. Por ello, en el presente experimento se estudian de una parte los resultados reproductivos de cerdas Retinto del Andévalo según orden de parto y estación de parto (primavera versus otoño) y, de otra, el peso al nacimiento, ganancia media diaria durante la lactación y peso al destete de los lechones según orden de parto, sexo y estación de nacimiento.
MATERIAL Y MÉTODOS
Se han controlado las variables reproductivas lechones nacidos totales (LNT), lechones nacidos vivos (LNV), lechones nacidos muertos (LNM), lechones muertos durante la lactación (LML) y lechones destetados (LD) de 250 camadas de la línea Retinto del Andévalo en la finca “Huerto Ramírez” de la Diputación de Huelva. Las cerdas se explotaron en cercas al aire libre con tiendas camping protectoras en gestación, lactación y cubrición. Durante la gestación las cerdas recibieron 2,2kg de pienso/día con 2950kcal de EM/kg, 13% de proteína bruta (PB) y el 0,54% de lisina. A las cerdas lactantes se les suministraron 3,3kg/día con 3100kcal de EM/kg, 16% de PB y el 0,82% de lisina Los lechones recibieron, después de la primera semana de edad, un pienso de pre arranque aportado ad libitum con 3250kcal de EM/kg, 19% de PB y el 1,50% de lisina. De otra parte, se controlaron el peso al nacimiento (PN), ganancia media diaria (GMD) durante la lactación, peso al destete (PD) y duración de la lactación (DL) de 164 lechones integrantes de 26 camadas de primero a cuarto parto. Los datos recabados fueron estudiados mediante análisis de varianza, covarianza, correlación y regresión simple y múltiple. Para las variables reproductivas los factores considerados fueron el orden de parto y la estación de parto, y su interacción incluyéndose en el modelo estadístico como covariables los LNT para la variable LNM, y LNV para las variables LML y LD. Para el PN, GMD y PD de los lechones los factores considerados fueron el orden de parto, sexo y estación de nacimiento (primavera vs otoño) incluyendo en el modelo estadístico correspondiente como covariables el PN los lechones amamantados (LA), calculados en función de los días presentes de lactación, y la duración de la lactación (DL). Por regresión y correlación se relacionaron diversas variables reproductivas y de crecimiento. Todos los análisis fueron realizados mediante el paquete estadístico SPSS 20.
RESULTADOS Y DISCUSIÓN
El orden de parto afectó significativamente (P<0,05) a los LNT, LNM, LML y LD (Tabla 1). La media de LNT y de LNV fue de 9,29 lechones y 6,66 lechones respectivamente siendo el valor observado de LNT superior al obtenido por Vázquez et al (1994) y al referenciado por Silió et al (2001) en las estirpes Guadyerbas y Torbiscal, y por Benito et al (1987) en la línea Valdesequera. Vázquez et al (1994) y Piñeiro et al (2012) en cerdas Ibéricas encontraron valores de LNV superiores a los evidenciados en este experimento. La prolificidad en la cerda Ibérica parece estar más influenciada por las pérdidas embrionarias al principio de la gestación que por el nivel de ovulación dándose una relación positiva entre esta variable y la mortalidad embrionaria (González-Bulnes et al., 2012). Puede que las cifras elevadas de prolificidad detectadas en este trabajo deriven, al margen de consideraciones genéticas, de que las cerdas de nuestro trabajo estuvieron teóricamente sometidas a un estrés discreto durante la gestación al haber transcurrido dicha fase fisiológica en cercas con casetas protectoras.
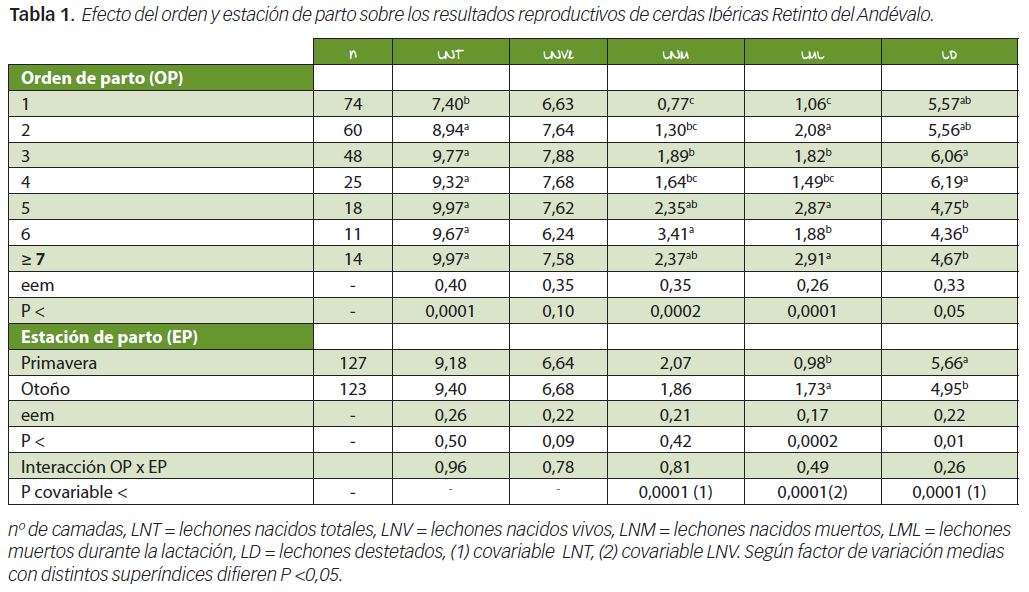
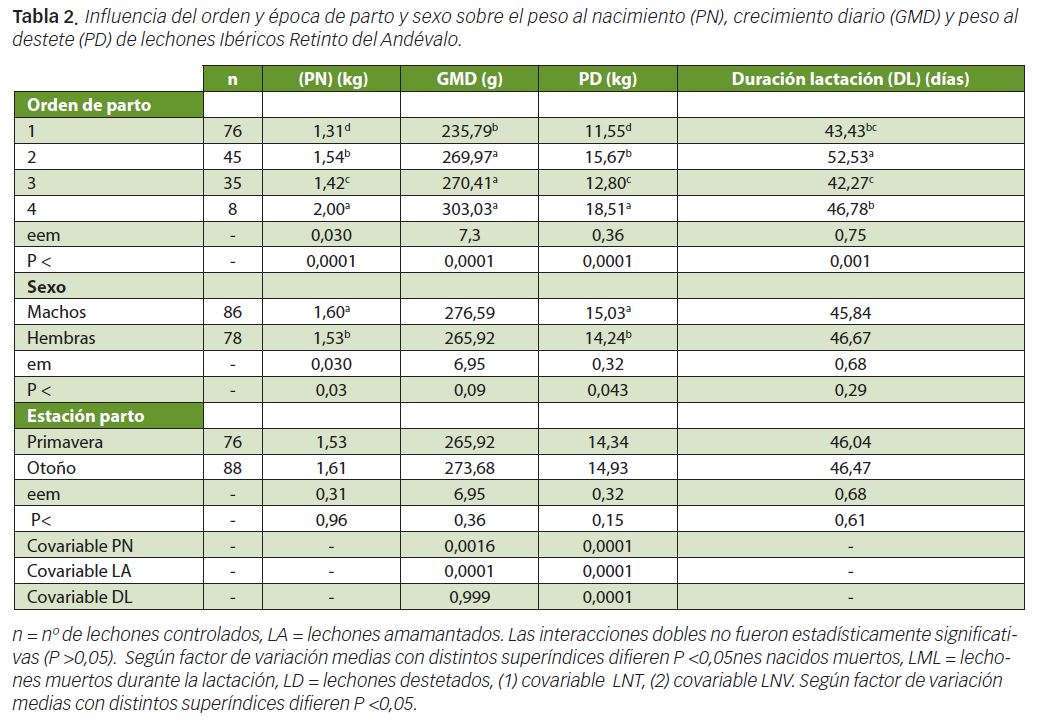
La superioridad de los LNT y LNV de cerdas multíparas versus primíparas también fue encontrada por Silió et al (2001) y Piñeiro et al (2012) obteniéndose los valores máximos de dichas variables reproductivas, como en nuestro estudio, entre el 3º y 7º parto. El valor de LNM también fue significativamente superior en cerdas multíparas que en primíparas alcanzándose los valores aritméticos más altos desde el 5º hasta el 7º parto. En la Tabla 3 puede observarse como los LNM aumentaron significativamente con los LNT.
Cuando en el modelo estadístico se introdujo la variable LNT como covariable los LNM desde el 1º al 7º o más partos fueron: 1,17, 1,27, 1,63, 1,51, 2,04, 3,19 y 2,07, resultados estadísticamente similares a los consignados en la Tabla 1. El aumento de la mortalidad al nacimiento con la prolificidad también ha sido observado por Daza et al (1993, 1999) aunque algunos autores como Piñeiro et al (2012) detectaron, en cerdas Ibéricas, porcentajes de mortalidad superiores en el 1º y 2º parto que en el 3º, 4º y 5º, probablemente debido a problemas distócicos.
La variable LML fue superior (P<0,05) en cerdas multíparas que en primíparas (Tabla 1) habiéndose detectado una relación positiva y significativa de esta variable con los LNV (Tabla 3), lo que se tradujo en que, en nuestro experimento, el número de LD fuera significativamente inferior (P <0,05) en los partos 5º, 6º y 7º o más que en el 3º y 4º, resultados que concuerdan los evidenciados por Piñeiro et al (2012). En la Tabla 3 puede observarse como los LD y los LNV estuvieron relacionados mediante una función cuadrática de estructura y = a + bx – cx2, lo que significa que los LD aumentaban con los nacidos vivos, pero a posteriori, en partos con elevada prolificidad disminuían debido al incremento de la mortalidad de lechones durante la lactación. Estos resultados concuerdan con los obtenidos por Daza y Gutiérrez–Barquín (1993) en cerdos blancos mejorados.
En lo concerniente al efecto de la estación de parto sobre las variables reproductivas y de mortalidad de lechones, en nuestro estudio, entre partos de primavera y otoño no se encontraron diferencias entre las variables LNT, LNV y LNM según estación, aunque debido a que la mortalidad de lechones durante la lactación fue superior en otoño, en esta estación se obtuvo un significativo (P<0,05) menor número de LD que en primavera. La influencia de la estación de parto sobre las variables reproductivas de cerdas Ibéricas se ha prestado a controversia. Así, Vázquez et al (1994) obtuvieron la prolificidad máxima en verano no observando diferencias entre primavera y otoño, mientras Silió et al (2001) referenciaron una mayor prolificidad en invierno. Benito et al (1987) obtuvieron más LNV y LD en primavera que en otoño, aunque la interacción año por estación de parto determinaba que tal resultado no se cumplía todos los años.
En lo que respecta a la mortalidad de lechones durante la lactación, la mayoría de los experimentos al respecto han observado mayor mortalidad en otoño-invierno que en primavera–verano. Las bajas temperaturas se traducen en el síndrome frío-hambre aplastamiento, y una reducción de la ingestión de calostro por el lechón determina un nivel inmunitario deficiente de efectos positivos sobre la mortalidad durante la lactancia (English y Morríson, 1985). De los resultados de la Tabla 3 puede colegirse que, conjuntamente, las variables LNV y LML explicaron, en nuestro trabajo, el 96% de la variabilidad de los LD.
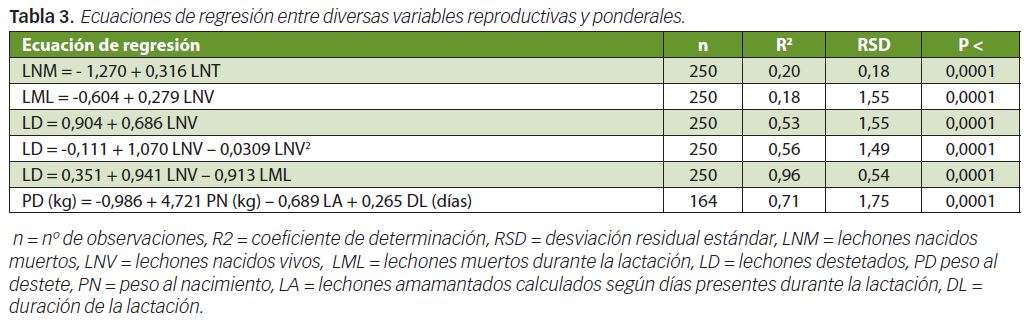
En la Tabla 2 reflejamos el efecto del orden de parto, sexo y estación de parto sobre el PN y GMD de los lechones durante la lactación. El PN del lechón aumentó con el orden de parto, era mayor en machos que en hembras y en dicha variable no influyó la estación de parto (primavera versus otoño). La GMD durante la lactación y el PD fue superior en los lechones de cerdas multíparas que en los de primíparas siendo el PD, dentro de las multíparas, más elevado en las cerdas de 4º parto que en las de 2º y 3º. Los machos tendieron (P<0,10) a crecer más que las hembras, y, al destete, los machos pesaron significativamente (P<0,05) más que las hembras Según puede observarse en la Tabla 3 el PD estuvo relacionado positivamente con el PN y con la DL y negativamente con LA, de modo que estas 3 precitadas variables, conjuntamente, explicaron el 71% de la variabilidad del PD.
Nuestros resultados de PN y de GMD obtenidos están de acuerdo con los observados en lechones Ibéricos por Blanco y Daza (2011) y fueron superiores a los encontrados en el metaanálisis realizado por Sánchez Esquiliche y Rodríguez Estévez (2011) y a los detectados por Benito et al (1987) en la línea Valdesequera. Nuestros PN elevados pueden explicarse por la baja prolificidad de las madres estudiadas para esta variable (7,90 LNT) y porque los animales fueron pesados dentro de las 24 horas siguientes al parto y no inmediatamente después del mismo, mientras que la elevada GMD obtenida se debió al bajo número de LA.
La confirmación de las anteriores ideas lo corrobora el que los coeficientes de correlación obtenidos entre el PN y los LNT fue de r = -0,47 (P<0,0001) y entre la GMD y LA r = -0,54 (P<0,0001). Según Benito et al (1995) (citado por Daza 2001) el PD a los 56 días fue ligeramente superior en machos que en hembras observándose los pesos más bajos en los LD en verano, mientras que, en nuestro experimento, la estación de parto (primavera frente a otoño) no tuvo influencia estadísticamente significativa sobre el PN, GMD y PD. En cerdos blancos mejorados el PN fue superior en machos que, en hembras, pero durante la lactancia, la GMD entre sexos fue similar (Daza et al., 1999 b).
CONCLUSIONES
Las cerdas Ibéricas de la línea Retinto del Andévalo exhiben una elevada prolificidad que debería continuarse estudiando por investigadores y técnicos, aunque los resultados al destete no son satisfactorios debido a la elevada mortalidad de lechones al nacimiento y durante la lactación inherentes a su modelo de explotación extensivo en camping. Por lo tanto, se sugiere cambiar el modelo de explotación hacia fórmulas más intensivas y a establecer estrategias adecuadas de manejo, alimentación, alojamiento y sanidad durante los periodos de gestación y lactación que reduzcan la mortalidad de lechones. De otra parte, en lo que se refiere a las variables PN, GMD durante la lactación y PD los valores evidenciados en este experimento se encuentran dentro del intervalo convencional o son superiores a los observados en otros estudios realizados con otras estirpes de cerdo Ibérico.
AUTORES
- Forero F.J., Venegas M. Diputación de Huelva.
- Daza A. Departamento de Producción Agraria (Unidad de Producción Animal). ETS de Ingeniería Agronómica, Alimentaria y de Biosistemas. Universidad Politécnica de Madrid (UPM).
Deja un comentario